Объемная скорость кровотока и метаболизм липидных соединений в стенке органов пищеварительного тракта коров в раннюю фазу лактации

Автор: Алиев А.А.
Журнал: Сельскохозяйственная биология @agrobiology
Рубрика: Обмен веществ и продуктивность
Статья в выпуске: 2 т.38, 2003 года.
Бесплатный доступ
На основе использования разработанных автором оперативных методов у коров черно-пестрой породы в раннюю фазу лактации оценивали объемную скорость кровотока печени и метаболизм липидов, липопротеинов и высокомолекулярных жирных кислот в стенке органов пищеварительной системы. Рассматриваются процессы циркуляции и рециркуляции липидов и их соединений в кровотоке.
Короткий адрес: https://sciup.org/142132875
IDR: 142132875 | УДК: 636.237.21:591.133.2
Volume blood velocity and metabolism of lipidic compounds in wall of organs of digestive tract in cows during early phase of lactation
On the basis of operative methods developed by the author in cows of the Black-and-White breed during early phase of lactation the volume velocity of liver bloodstream and metabolism of lipids, lipoproteins and high molecular fatty acids in wall of digestive system organs were estimated. The circulation processes and recirculation of lipids and their compounds in blood are considered. It was shown, that absorbtion and transport of metabolites occur simultaneously with complex interactions of metabolic processes result in the formation of blood plasm of portal vein, and further correction of metabolic status of plasm in total circulation occurs in the liver.
Текст научной статьи Объемная скорость кровотока и метаболизм липидных соединений в стенке органов пищеварительного тракта коров в раннюю фазу лактации
Известно, что доминирующая роль при синтезе молока и его компонентов принадлежит процессам, происходящим в пищеварительном канале и печени (1, 2). Задачей настоящей работы было определение количественных показателей притока и оттока крови через стенки органов пищеварительного тракта и оценка влияния этих процессов на метаболизм липидов, высокомолекулярных жирных кислот (ВМЖК) и липопротеинов (ЛП) у коров черно-пестрой породы в раннюю фазу лактации.
Методика. Объектом исследования служили коровы черно-пестрой породы ( n = 3), удои которых за предыдущую лактацию составляли 5000 кг молока. Животным скармливали зимний сбалансированный рацион, включающий комбикорм, сено, подсолнечниковый шрот, разнотравный силос и кормовую свеклу (3). Через 2 нед после отела при среднесуточном удое 20,7 ± 0,72 кг молока коровам были имплантированы катетеры в плечеголовной ствол аорты, а также портальную, общую брыжеечную и печеночные вены (4, 5). Кровь из катетеризованных сосудов брали до и через 3 ч после утреннего кормления. В образцах кормов определяли содержание сухого вещества; в крови — концентрацию общих липидов (6), соединений липидов, ЛП и хиломикронов (ХМ), а также ВМЖК (7). Объемную скорость кровотока оценивали методом индикаторного разведения при инфузии п -аминогиппуровой кислоты в общую брыжеечную вену (5). Гидратацию портальной крови рассчитывали по величине гемоглобинового индекса (4). Статистическую обработку данных проводили по Стьюденту.
Результаты. У коров живой массой 480 ± 0,25 кг при потреблении 13,7 ± 0,6 кг сухого вещества и суточном удое 20,4 ± 1,7 кг молока (продукция молочного жира и белка — соответственно 706,0 ± 0,93 и 533,0 ± 0,68 г/сут) объем крови, оттекающей от желудочно-кишечного канала и притекающей по воротной вене к печени, составлял 33,9 ± 2,8 л/мин на 1 кг живой массы, а объем крови, оттекающей от печени в заднюю полую вену, — 42,0 ± 3,30 л/мин. Через 3 ч после утреннего кормления объем притекающей в желудочно-кишечный канал артериальной крови достигал 68,08, а оттекающей — 72,13 мл/мин на 1 кг живой массы; объем оттекающей от печени крови — 89,36 мл/мин. Б о льший объем крови, оттекающей от печени в заднюю полую вену, объясняется двойным притоком крови к печени — по воротной вене и по печеночной артерии. При этом объем притекающей к печени по печеночной артерии крови составлял 15 % от объема портальной крови (4).
В образцах крови, полученных из катетеризированных сосудов, преобладали эфиры холестерола, триацилглицеролы и фосфолипиды — соответственно 33,42; 30,38 и 8,20 % от общего количества липидов. До кормления животных концентрация общих липидов в артериальной крови была максимальной, а в воротной вене — ниже на 16,85 мг% (или на 5,39 %), то есть артериовенозная разница была положительной (табл. 1). Следовательно, в пищеварительном канале происходило всасывание из притекающей крови общих липидов и их соединений, а именно: моно- и диацилгли-церолов — 0,95, неэтерифицированных жирных кислот (НЭЖК) — 6,79, эфиров хо- лестерола — 8,49 мг%. В то же время в кровь воротной вены через стенки пищеварительного канала выделялось 4,53 и 1,95 мг% соответственно фосфолипидов и свободного холестерола.
-
1. Концентрация липидных соединений в крови различных сосудов желудочнокишечного канала коров черно-пестрой породы в зависимости от времени взятия пробы (мг%)
-
2. Концентрация липидных соединений в плазме крови кровотока печени коров черно-пестрой породы в зависимости от времени взятия пробы (мг%)
|
Липидные соединения До кормления |
Через 3 ч после кормления |
|
|
А р т е р и а л ь н а я к р о в ь |
||
|
Общие липиды |
312,50 ± 29,35 |
301,23 ± 27,13 |
|
Фосфолипиды |
27,54 ± 2,29 |
31,49 ± 4,48 |
|
Моно-, диацилглицеролы |
7,14 ± 1,09 |
10,86 ± 1,56 |
|
Холестерол свободный |
6,42 ± 1,12 |
14,15 ± 12,91 |
|
НЭЖК |
28,71 ± 3,60 |
27,84 ± 5,53 |
|
Триацилглицеролы |
115,50 ± 10,24 |
92,18 ± 10,16 |
|
Эфиры холестерола |
127,38 ± 22,19 |
124,71 ± 20,88 |
|
В е н о з н а я к р о в ь (воротная вена) |
||
|
Общие липиды |
295,65 ± 19,20 |
315,68 ± 20,67 |
|
Фосфолипиды |
32,07 ± 5,91 |
27,77 ± 3,60 |
|
Моно-, диацилглицеролы |
6,19 ± 0,77 |
8,94 ± 1,52 |
|
Холестерол свободный |
8,37 ± 2,66 |
9,78 ± 1,10 |
|
НЭЖК |
21,92 ± 2,91 |
28,46 ± 3,84 |
|
Триацилглицеролы |
108,21 ± 12,37 |
117,00 ± 16,43 |
|
Эфиры холестерола |
118,89 ± 12,90 |
123,73 ± 18,23 |
|
А р т е р и о в е н о з н а я р а з н и ц а |
||
|
Общие липиды |
16,85 |
–14,45 |
|
Фосфолипиды |
–4,53 |
3,72 |
|
Моно-, диацилглицеролы |
0,95 |
1,92 |
|
Холестерол свободный |
–1,95 |
4,37 |
|
НЭЖК |
6,79 |
–0,62 |
|
Триацилглицеролы |
7,30 |
–24,82 |
|
Эфиры холестерола |
8,49 |
0,98 |
П р и м е ч а н и е. НЭЖК — неэтерифицированные жирные кислоты.
Как известно, у жвачных в отличие от моногастричных животных при нормальных условиях кормления не удается создать «голодный фон» пищеварения, поэтому у первых нет доминирующего времени всасывания, а происходит непрерывный обмен веществ между кровью и стенками органов пищеварительного тракта (1). Это положение подтверждается полученными нами данными. Через 3 ч после кормления через стенку пищеварительного канала в портальную кровь с учетом объемной скорости тока за 1 мин выделялось 43,8 мкг общих липидов на 1 кг живой массы коровы, в том числе НЭЖК, триацилглицеролов и эфиров холестерола — соответственно 4,0; 24,8 и 15,0 мкг. При этом одновременно из артериальной крови поглощались фосфолипиды, монодиацилглицеролы и свободный холестерол — соответственно 3,7; 2,0 и 4,37 мкг/мин на 1 кг живой массы.
До кормления животных концентрация общих липидов в плазме крови, притекающей к печени, была выше, чем в оттекающей. Разница по кровотоку между притекающей и оттекающей кровью (П–О) была положительной — 27,59 мг%, то есть печень поглощала из притекающей крови общие липиды, в том числе триацилг-лицеролы и эфиры холестерола: П–О соответственно 16,48 и 27,79 мг% (табл. 2). Одновременно из печени в
Липидные соединения До кормления Через 3 ч после кормления
П р и т е к а ю щ а я к р о в ь (портальная вена)
|
Общие липиды |
301,50 ± 12,40 |
312,50 ± 17,60 |
|
Фосфолипиды |
31,16 ± 1,82 |
28,52 ± 1,81 |
|
Моно-, диацилглицеролы |
6,31 ± 0,86 |
9,33 ± 1,88 |
|
Холестерол свободный |
8,22 ± 1,45 |
10,65 ± 1,51 |
|
НЭЖК |
23,27 ± 2,74 |
28,32 ± 2,62 |
|
Триацилглицеролы |
111,80 ± 11,42 |
112,04 ± 11,92 |
|
Эфиры холестерола |
120,74 ± 13,82 |
123,94 ± 11,33 |
О т т е к а ю щ а я к р о в ь
Через 3 ч после кормления в портальной крови было отмечено незначительное повышение концентрации общих липидов, свободного холестерола и НЭЖК, а также снижение концентрации фосфолипидов — с 31,16 до 28,52 мг%; в оттекающей от печени крови — достоверное увеличение концентрации общих липидов (Р < 0,05), фосфолипидов (Р < 0,001), моно- и диацилглицеролов. При этом показатель П–О был отрицательным, то есть в печени происходил активный метаболизм этих соединений. По классу триацилглицеролов и эфиров холестерола разница П–О составляла соответственно 1,20 и 14,48 мг% (см. табл. 2).
С учетом объемной скорости кровотока мы установили, что за 1 мин в печени было синтезировано общих липидов, фосфолипидов, моно- и диацилглицеролов, свободного холестерола, НЭЖК соответственно 57,5; 30,5; 12,0; 8,4; 6,6 мкг/кг живой массы. В то же время отмечено всасывание триацилглицеролов и эфиров холестерола из притекающей к печени крови — соответственно 10,3 и 15,7 мкг/мин на 1 кг живой массы.
Хиломикроны — наиболее легкая фракция липопротеинов (плотность < 1,00 г ⋅ см–3), образующихся в процессе всасывания липидов в эпителиоцитах тонкого кишечника. Концентрация ХМ в плазме крови из разных сосудов у лактирующих коров составляла 6,3-10,5 % от общего количества липидов. Сравнительно небольшое количество этой фракции в крови относительно лимфы (28,0-58,0 %) объясняется тем, что ХМ разрушаются под действием липопротеинлипазы, локализованной на поверхности эндотелия капилляров (2). Через 3 ч после кормления концентрация ХМ в плазме артериальной крови составляла 18,07 мг%, артериовенозная разница — 5,50 мг%, то есть с учетом объемной скорости кровотока за 1 мин через стенку пищеварительного канала в кровь выделялось 5,0 мкг/кг живой массы, а в печени синтезировалось 11,0 мкг/кг живой массы ХМ (рис., а).
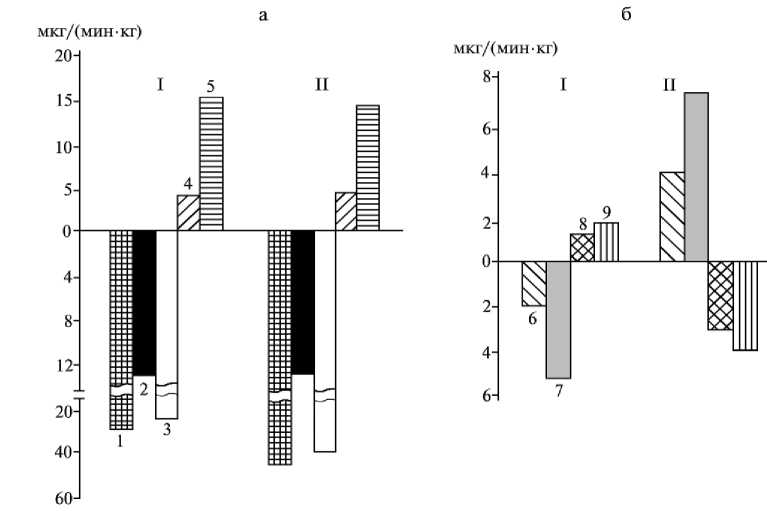
Объемная скорость кровотока липидных соединений (а) и высокомолекулярных жирных кислот (б) (мкг/мин на 1 кг живой массы) через стенки органов желудочно-кишечного тракта коров чернопестрой породы: I — артериовенозная разница, II — разница между притекающей к печени и оттекающей кровью; 1 — общие липиды; 2 — хиломикроны; 3, 4 и 5 — липопротеины соответственно высокой, низкой и очень низкой плотности; 6, 7, 8 и 9 — соответственно пальмитиновая (C 16:0 ), стеариновая (С 18:0 ), олеиновая (C 18:1 ) и линолевая (C 18:2 ) кислоты.
Фракция липопротеинов очень низкой плотности (ЛПОНП— 1,01-1,06 г ⋅ см– 3) составляла 50-70 % от общего количества липидов, а концентрация этих соединений в крови из всех сосудов — 200,0-250,3 мг%. Через 3 ч после кормления артериовенозная разница достигала 32,71 мг%; за 1 мин через стенку пищеварительного канала в кровь выделялось 34,0, а в печени синтезировалось 32,0 мкг/кг живой массы ЛПОНП. Концентрация фракций липопротеинов низкой (ЛПНП) и высокой (ЛПВП) плотности была значительно меньше, чем таковая ЛПОНП. При этом положительная артериовенозная разница свидетельствует об активном всасывании ЛПНП и ЛПВП (соответственно 14,90 и 9,57 мг%) из притекающей крови в стенке пищеварительного канала — соответственно 6,0 и 1,7 мкг/мин. Следовательно, эпителиоциты кишечника активно участвуют в синтезе ХМ и ЛПОНП, используя для трансформирования ЛПНП и ЛПВП. Скорость образования в печени ХМ и ЛПОНП составляла соответственно 11,0 и 52,0 мкг/мин на 1 кг живой массы, причем в процессе метаболизма использовались ЛПНП и ЛПВП.
В составе крови коров было идентифицировано 22 ВМЖК, среди которых доминирующими являлись пальмитиновая (С 16:0 ), стеариновая (C 18:0 ), олеиновая (С 18:1 ) и линолевая (С 18:2 ) кислоты. Через 3 ч после кормления концентрация насыщенных жирных кислот увеличивалась как в артериальной, так и портальной крови преимущественно за счет стеариновой кислоты, количество которой в артериальной крови возрастало в 1,2 раза. Большая артериовенозная разница свидетельствует об активном всасывании насыщенных жирных кислот из желудочно-кишечного канала. Через стенку кишечника в кровь выделялось 2,0 и 6,0 мкг/мин на 1 кг живой массы соответственно пальмитиновой и стеариновой кислот (рис., б).
После кормления в крови возрастала концентрация пента- и гептадеценовой кислот микробиального происхождения вследствие повышения активности процессов пищеварения. Концентрация короткоцепочечных жирных кислот (С6-С14) в артериальной и портальной крови до кормления составляла соответственно 1,38±0,07 и 1,11±0,27 мг%, а через 3 ч после кормления она увеличивалась в 1,2 и 1,4 раза. По концентрации ВМЖК в притекающей к печени и оттекающей крови до и после кормления выявлена существенная разница П–О. Положительные значения разницы П–О до кормления животных по пальмитиновой (C16:0) и стеариновой (C18:0) кислотам (соответственно 3,01 и 3,45 мг%), а также отрицательные — по олеиновой (C18:1) и линолевой (C18:2) свидетельствуют о притоке в печень с портальной кровью насыщенных жирных кислот и оттоке в общую циркуляцию крови ненасыщенных форм, что отражает десатуриазную активность гепатоцитов. Микробиальная гидрогенизация в рубце ненасыщенных жирных кислот, поступающих с липидами корма, существенно лимитирует потребность организма коров, в том числе и молочной железы, в ненасыщенных жирных кислотах, а поэтапная дегидрогенизация насыщенных жирных кислот, особенно стеариновой, в энтероцитах, гепатоцитах и секреторных клетках молочной железы обеспечивает эту потребность.
Таким образом, у коров живой массой 480 кг при потреблении 13,7 кг сухого вещества отток крови через стенки органов желудочно-кишечного тракта до утреннего кормления составляет 33,9 л/мин на 1 кг живой массы при положительной артериовенозной разнице, а через 3 ч после кормления объемная скорость кровотока возрастает в 2 раза при отрицательной артериовенозной разнице. До кормления концентрация общих липидов в артериальной крови оказывается выше, чем в портальной, при этом артериовенозная разница составляет 16 мг/кг живой массы, то есть в органах желудочно-кишечного канала происходит всасывание липидов и их соединений из артериальной крови. В то же время в кровь воротной вены поступают триацилгли-церолы и свободный холестерол. Через 3 ч после кормления характер липидного обмена меняется: в портальную кровь поступают общие липиды, неэтерифицированные жирные кислоты, триацилглицеролы и эфиры холестерола при одновременном поглощении из артериальной крови фосфолипидов, моноацилглицеролов и свободного холестерола. Через стенки органов пищеварительного канала в портальную кровь выделяются хиломикроны и липопротеины очень низкой плотности при одновременном всасывании из артериальной крови липопротеинов низкой и высокой плотности, то есть в синтезе этих соединений активно участвуют энтероциты. С учетом гидрота-ции и гематокрита крови нами установлено, что портальная кровь в зависимости от кормления животных то сгущается, то разжижается в течение суток с соответствующим увеличением или уменьшением числа форменных элементов в единице объема. Отсюда следует, что понятия всасывания и транспорта метаболитов в желудочнокишечном тракте следует рассматривать с новых позиций. Эти процессы протекают на фоне сложного взаимодействия, а также циркуляции и рециркуляции метаболитов между органами кровеносной и пищеварительной систем, результатом которых является образование плазмы крови, оттекающей в портальную систему (1, 2). При этом печень осуществляет дальнейшую коррекцию метаболического статуса плазмы крови на основе гормональной и нервной регуляции метаболических взаимодействий как между органами желудочно-кишечного тракта, так и всего организма.
Л И Т Е Р А Т У Р А
-
1. А л и е в А.А. Лимфа и лимфообращение у продуктивных животных. Л., 1982.
-
2. А л и е в А.А. Обмен веществ у жвачных животных. М., 1997.
-
3. К а л а ш н и к о в А.П., К л е й м е н о в Н. И., Щ е г л о в В.В и др. Нормы и рационы кормления
сельскохозяйственных животных. М., 1985.
-
4. А л и е в А.А. Оперативные методы исследования сельскохозяйственных животных. Л., 1974.
-
5. А л и е в А.А. Экспериментальная хирургия. М., 1998.
-
6. F o l c h E., L e e s M., S t a n l e y H.S. A simple method for isolation and purification of total lipids from animal tissues. J. Biol. Chem., 1975, 26: 497-509.
-
7. Методы биохимических исследований /Под ред. Б.Д. Кальницкого. Боровск, 1997.
Всероссийский НИИ физиологии, биохимии и Поступила в редакцию питания сельскохозяйственных животных, 7 февраля 2002 года
249000, Калужская обл., г. Боровск
VOLUME BLOOD VELOCITY AND METABOLISM OF LIPIDIC COMPOUNDS IN WALL OF ORGANS OF DIGESTIVE TRACT
IN COWS DURING EARLY PHASE OF LACTATION
S u m m a r y
On the basis of operative methods developed by the author in cows of the Black-and-White breed during early phase of lactation the volume velocity of liver bloodstream and metabolism of lipids, lipoproteins and high molecular fatty acids in wall of digestive system organs were estimated. The circulation processes and recirculation of lipids and their compounds in blood are considered. It was shown, that absorbtion and transport of metabolites occur simultaneously with complex interactions of metabolic processes result in the formation of blood plasm of portal vein, and further correction of metabolic status of plasm in total circulation occurs in the liver.
Новые книги
Б а б и н а М.П. Иммунная реакция цыплят-бройлеров в онтогенезе и ее коррекция микробными препаратами. Витебск: ВГАВМ, 2002, 114 с.
В монографии обобщены результаты изучения закономерностей формирования иммунного статуса цыплят-бройлеров в эмбриональный и постовариальный периоды онтогенеза по данным автора и источникам литературы. Показано влияние защитных факторов яйца на иммунную реакцию цыплят в зависимости от периода яйцекладки. Рассматривается структурная организация иммунной системы. Приведены постовариальные иммунологические и гематологические показатели цыплят-бройлеров, а также отмечены возрастные особенности гемопоэза. Представлена динамика клеточных и гуморальных факторов иммунной защиты в постовариальном онтогенезе. Отражена зависимость иммунного статуса цыплят от обмена веществ. Обсуждается эффективность новых пробиотиков и иммунокорректоров при стимуляции местной и системной защиты, профилактике иммунодефицитов и болезней молодняка птицы, в том числе желудочнокишечных заболеваний и гиповитаминозов.
А р ь к о в А.А., Г о р л о в И.Ф., Б е -л я е в А.И. и др. Бишофит и другие минеральные вещества в птицеводстве. Волгоград: типогр. Химпром, 2002, 180 с.
В монографии освещены вопросы использования местных минеральных источников природного бишофита, донских известняков, эльтонской соли, а также премикса из этих веществ в качестве добавок в рационы кур и цыплят-бройлеров. Приведена экономическая оценка эффективности применения минеральных источников Волгоградского региона. Отмечено, что бишофит является комплексным соединением макроэлемента магния и значительного числа (до 70) микроэлементов, применение которого оказывает положительное влияние на яйценоскость кур, качество скорлупы яиц, стимулирует рост и мясную продуктивность цыплят-бройлеров, укрепляет костяк птицы, уменьшая отход за период откорма. При этом значительно снижаются затраты корма. Показаны биогенные свойства природного бишофита при обработке инкубационных яиц. Даны рекомендации по дозам минеральных веществ при добавлении в рационы и обработке инкубационных яиц.
