Оценка генетического полиморфизма сортов, линий и мутантов гороха посевного (Pisum sativum L.) с помощью ДНК-маркеров на основе ретротранспозонов
 с помощью ДНК-маркеров на основе ретротранспозонов")
Автор: Кокаева З.Г., Гостимский С.А.
Журнал: Сельскохозяйственная биология @agrobiology
Рубрика: ДНК-технологии в селекции растений
Статья в выпуске: 3 т.42, 2007 года.
Бесплатный доступ
Резюме. IRAP-метод использовали для изучения инсерционного полиморфизма ретротранспозо-нов группы Ту1-copia у 16 сортов, линий и мутантов гороха посевного (Pisum sativum L.). Оценили эффективность десяти комбинаций праймеров при выявлении полиморфизма у анализируемых образцов, получили полиморфные маркеры генома гороха. На основании IRAP-анализа определили генетические дистанции и построили дендрограмму, отражающую генетическое сходство и родство между исследованными образцами. Для каждого из анализируемых сортов и линий получили индивидуальный IRAP-спектр с целью выявления сортоспецифичных фрагментов, пригодных для паспортизации и маркирования изученных форм.
Горох, филогенетический анализ, ретротранспозон
Короткий адрес: https://sciup.org/142134816
IDR: 142134816 | УДК: 635.656:575.17.015.3:577.088
Estimation of genetic polymorphism in cultivars, lines and mutants of garden pea (Pisum sativum L.) using DNA-markers on the basis of retrotransposons
IRAP-method was used for study of insertional polymorphism of Ту1-copia retrotrans-posons in 16 cultivars, lines and mutants of garden pea (Pisum sativum L.). 10 combinations of primers distinguishing polymorphism in analyzed lines were designed. 72 polymorphic markers were found. Using IRAP-analysis genetic distances were counted and comparative dendrogramm reflexing genetic similarity and relationship between samples analyzed. Individual IRAP-spectrums were analyzed for every cultivar and line; some cultivar-specific fragments suitable for identification and marking of analyzed forms were found. Usage of IRAP-analysis for such purposes in pea cultivars and lines was shown may be prospective.
Текст краткого сообщения Оценка генетического полиморфизма сортов, линий и мутантов гороха посевного (Pisum sativum L.) с помощью ДНК-маркеров на основе ретротранспозонов
Ранее для паспортизации и идентификации сортов и линий гороха посевного мы применили RAPD-метод (9).
Целью настоящей работы было изучение генетического полиморфизма сортов, линий и мутантов гороха Pisum sativim L. с использованием ДНК-маркеров на основе ретротранспозонов группы Ty1- copia .
Методика . Анализировали растения 16 образцов гороха посевного: сорта Демон, Филби, Флагман, Виола, Немчиновский, Ранний Зеленый, Торсдаг, Finale, Rondo из коллекции кафедры генетики биологического факультета МГУ им. М.В. Ломоносова, линии L-131, L-851, L-1132, L-1238, L-102 из Института генетики сельскохозяйственных растений (Швеция) и хлорофильные мутанты Хл 42 и Хл 15, полученные обработкой семян соответственно сортов Немчиновский и Торсдаг этилметансульфона-том. Тотальную ДНК выделяли из молодых листьев методом CTAB с изменениями (10).
В работе использовали следующие праймеры (11):
Tps 3 5'-CCTTTGGGATATTAACCACAC-3'
Tps 6 5'-GTGAGATAGTTATATGTC-3'
Tps 7 5'-CTATAATACATAACAAGC-3'
Tps 10 5'-GGAATGATAGGCCTTGCC-3'
*RNaseH 1 5'-MGNACNAARCAYATHGA-3'
*RNaseH 2 5'-GCNGAYATNYTNACNAA-3'
* Последовательности вырожденных праймеров.
Амплификацию проводили в термоциклере «Терцик» («ДНК-технология», Россия) по следующей программе: предварительная денатурация при 95 оС в течение 2 мин; 30 циклов при 95 оС в течение 60 с; при температуре отжига в течение 60 с; при 72 оС в течение 2 мин; конечная элонгация при 72 оС в течение 10 мин. Реакционная смесь объемом 25 мкл содержала 50 нг ДНК, 2 ед. Taq-полимеразы («Силекс М», Россия), 2,5 мкл стандартного 10-кратного буфера для ПЦР («Силекс М», Россия), 25 пМ каждого праймера, 2,5 мМ Mg2+ и 0,25 мМ dNTP; на смесь наслаивали две капли минерального масла.
Работа выполнена при финансовой поддержке РФФИ (грант № 07-04-00652) и программы президиума РАН «Динамика генофондов растений, животных и человека».
Продукты амплификации разделяли электрофорезом в 2 % агарозном геле с однократным ТВЕ-буфером и окрашивали бромистым этидием. Длину фрагментов определяли по маркерам молекулярной массы 100 bp + 1,5+3 Kb DNA Ladder («Сибэнзим», Россия). Гели просматривали в УФ-свете ( λ = 312 нм); результаты документировали с использованием цифровой фотокамеры. Э ксперименты проводили в нескольких повторностях.
Для количественной оценки полиморфизма и определения генетического расстояния между исследованными формами данные IRAP-анализа были представлены в виде матрицы состояний, в которой наличие или отсутствие в IRAP-спектрах фрагментов одинаковой длины обозначили соответственно 1 или 0. По матрице состояний была составлена матрица различий, генетические расстояния в которой рассчитывались по формуле Nei и Li (12) с помощью компьютерной программы Treecon (13).
На основании полученной матрицы невзвешенным парно-групповым методом (UPGMA — Unweighted Pair-Group Method) (13) была построена генеалогическая дендрограмма, отражающая степень различий между IRAP-спектрами исследуемых форм. Для построения дендрограммы использовали компьютерную программу Treecon (13). Оценку значимости генеалогических реконструкций проводили методом бутстрепа (14). Топология дерева в участках со значениями больше 50 % может считаться надежно установленной.
Результаты . Из 21 протестированной комбинации праймеров десять эффективно выявляли полиморфизм, а три могли быть использованы при паспортизации сортов и линий гороха (табл. 1). Для оценки полиморфизма генома в каждой комбинации праймеров определяли отношение числа полиморфных фрагментов к общему числу амплифицированных. Было показано, что у проанализированных сортов, линий и мутантов гороха отдельные праймеры выявляли от 7 до 25 фрагментов размером 1501500 п.н., из которых от 4 до 13 были полиморфными (рис. 1). Всего получили 130 ДНК-фрагментов, из них 72 были полиморфными. Общий уровень полиморфизма с десятью комбинациями IRAP-праймеров составил 55,38 %. При оценке внутрилинейной изменчивости у линии L-131 показали, что IRAP-спектры растений гороха, полученные с парой праймеров (Tps 7 - Tps 10), идентичны (рис. 2).
1. Эффективность комбинаций праймеров при выявлении полиморфизма IRAP-фрагментов ДНК
|
Праймер Праймер |
Tps 3 |
Tps 6 |
Tps 7 |
Tps 10 |
RNaseH 1 |
RNaseH 2 |
Для количественной оценки полиморфизма и определения степени дивергенции между изученными формами гороха данные, полученные при анализе IRAP-полиморфизма, представили в виде матрицы состояний 130 бинарных признаков, которую использовали для расчета генетических расстояний и построения дендрограммы.
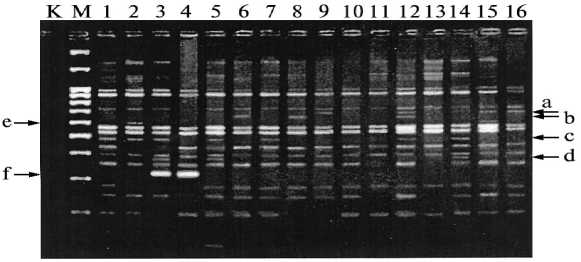
Рис. 1 . Продукты IRAP-амплификации ДНК сортов (1-7, 10, 11), линий (8, 9, 12, 15, 16) и мутантов гороха (13, 14) с парой праймеров (Tps 7 - Tps 10): 1 — Демон, 2 — Филби, 3 — Флагман, 4 — Виола, 5 — Немчиновский, 6 — Rondo, 7 — Finale, 8 — L-1238, 9 — L-102, 10 — Ранний Зеленый, 11 — Торсдаг, 12 — L-851, 13 — Хл 15, 14 — Хл 42, 15 — L-131, 16 — L-1132. К — отрицательный контроль; М — маркер молекулярной массы 100 bp + 1,5 + 3 Kb DNA Ladder. Стрелками отмечены полиморфные фрагменты: a — 690 п.н., b — 680 п.н., с — 490 п.н., d — 380 п.н., e — 600 п.н., f — 320 п.н.
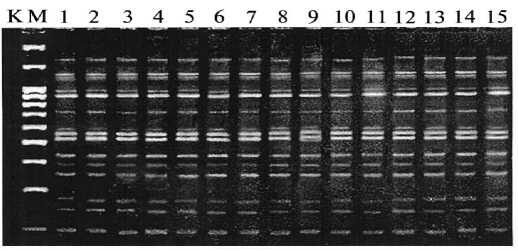
Рис. 2. IRAP-спектры ДНК-фрагментов у растений гороха линии L-131, полученные с парой праймеров ( Tps 7 - Tps 10): 1-15 — индивидуальные растения гороха линии L-131. К — отрицательный контроль; М — маркер молекулярной массы 100 bp + 1,5 + 3 Kb DNA Ladder.
Уровень различий между мутантами Хл 42, Хл 15 и исходными сортами составил 0 %, средний уровень полиморфизма между формами — 23,74 %, что согласуется с ранее полученными данными (9).
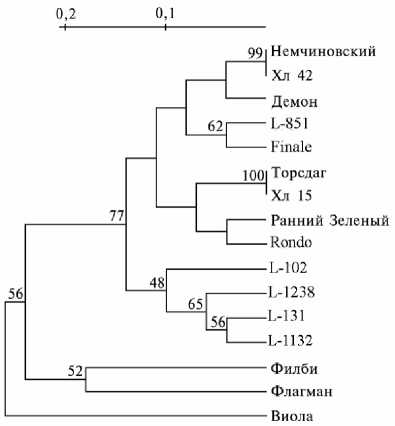
Рис. 3. Дендрограмма исследованных линий, сортов и мутантов гороха (по методу UPGMA). Верхняя шкала — генетическое расстояние по Nei и Li (12). Цифрами указаны значения бут-стрепа, %.
Дендрограмма, отражающая степень различий между IRAP-спектрами образцов гороха, представлена на рисунке 3. Исходя из матрицы различий, наибольшие генетические расстояния были установлены между сортом Флагман и маркерными линиями (37,77 %), наименьшие — между мутантами и исходными сортами (0 %). Дендрограмма показывает, что сорт Немчиновский и полученный из него мутант Хл 42, а также сорт Демон кластеризуются в одну группу; сорт Торсдаг и полученный из него мутант Хл 15, сорта Ранний Зеленый и Rondo — в другую (уровень различий между кластерами составил 0,03). Вместе кластеризуются маркерные линии шведской селекции L-1132, L-131, L-1238 и линия L-102 (уровень различий между маркерными линиями и сортами — 0,02-0,1). Сорт Виола оказался обособленным от остальных, что, возможно, объясняется их разным происхождением. Эти результаты также согласуются с полученными нами ранее (9).
-
2. Матрица ДНК-фрагментов для паспортизации сортов, линий и мутантов гороха посевного ( Pisum sativum L.) с помощью IRAP- маркеров
Образец
Длина ДНК-фрагментов, п.н.
710
1 690
680
600 490
440
380
320
280
250
200
Демон
+
–
–
Сорта – +
+
–
–
+
+
+
Филби
+
–
–
+ +
–
–
–
–
+
+
Флагман
+
–
–
– +
–
+
+
–
+
-
Виола
–
–
–
– –
+
–
+
–
–
+
Немчиновский
+
–
–
– +
+
+
–
+
+
+
Rondo
+
–
+
– –
+
–
–
+
+
+
Finale
+
+
–
– +
+
–
–
+
+
+
Ранний Зеленый
+
–
–
–
+
–
–
+
+
+
Торсдаг
+
–
–
– –
+
–
–
+
–
+
L-851
+
+
+
Линии – +
+
–
–
+
+
+
L-131
+
+
–
+ –
+
–
–
+
+
+
L-1132
+
+
+
+ –
+
–
–
+
+
+
L-1238
+
+
+
+ –
+
–
–
+
+
–
L-102
+
+
–
– –
+
–
–
+
+
–
Хл 15
+
–
Му –
тантные формы – – +
–
–
+
–
+
Хл 42
+
–
–
– +
+
+
–
+
+
+
П р и м е ч а н и
е. «+»
и «–»
означает
наличие и отсутствие
маркера.
ДНК-фрагменты
получены с
комбинацией праймеров (Tps 7 - Tps 10).
Для однозначной идентификации изученных форм гороха посевного методом IRAP-PCR оказалось достаточно использовать несколько пар праймеров: (Tps 7 - Tps 10), (Tps 3 - Tps 6), (Tps 3 - Tps 7). IRAP-маркирование с этими праймерами позволяло амплифицировать уникальные для каждого образца IRAP-спектры у всех исследованных сортов и линий. Так, в спектре, полученном с комбинацией праймеров (Tps7 - Tps 10), фрагмент размером 680 п.н. присутствовал только у сорта Rondo и трех маркерных линий — L-1238, L-102, L-1132; фрагмент 690 п.н. — только у маркерных линий (кроме L-1238); фрагмент 600 п.н. обнаружили у сорта Филби и у двух маркерных линий — L-1238, L-1132; фрагмент 320 п.н. — только у сортов Флагман и Виола, а фрагмент 380 п.н. — у сортов Флагман и Немчиновский, а также у мутанта сорта Немчиновский Хл 42 (см. рис. 1). Фрагмент 280 п.н. отсутствовал только у трех сортов — Филби, Флагман и Виола.
Выявленная специфичность спектров позволяет проводить быструю идентификацию образцов. Набор ДНК-фрагментов, полученный в результате амплификации, может служить молекулярногенетическим паспортом сорта или линии гороха (табл. 2).
Таким образом, использование IRAP-метода позволяет достаточно эффективно дифференцировать разные генетические формы гороха. Полученные IRAP- маркеры можно использовать не только для идентификации и паспортизации сортов и линий гороха, но также для филогенетических исследований и в работе по картированию хромосом. По сравнению с другими маркерными системами выявления генетического полиморфизма IRAP-метод чувствительнее и специфичнее, при этом он прост в исполнении, не требует использования радиоактивных изотопов и эндонуклеаз рестрикции, полученные результаты хорошо воспроизводимы. Единственное ограничение применения метода состоит в том, что необходимо знать нуклеотидную последовательность терминальных участков мобильных элементов.
Л И Т Е Р А Т У Р А
-
1. Г о с т и м с к и й С.А., К о к а е в а З.Г., К о н о в а л о в Ф.А. Изучение организации и изменчивости генома растений с помощью молекулярных маркеров. Генетика, 2005, 41, 4: 1-15.
-
2. P e a r c e S.R., K n o x M., E l l i s T.H.N. e.a. Pea Ty1- copia group retrotransposons: transpositional activity and use as markers to study genetic diversity in Pisum . Mol. Gen. Genet., 2000, 263: 898-907.
-
3. K u m a r A., B e n n e t z e n J. Plant retrotransposons. Annu. Rev. Genet., 1999, 33: 479-532.
-
4. T a t o u t C., W a r w i c k S., L e n o i r A. e.a. Sine insertions as clad markers for wild Crucifer species. Mol. Biol. Evol., 1999,
16: 1614-1621.
-
5. W a u g h R., M c L e a n K., F l a v e l l A.J. e.a. Genetic distribution of BARE-1-like retrotransposable elements in the barley genome revealed by sequence-specific amplification polymorphisms (S-SAP). Mol. Gen. Genet., 1997, 253: 687-694.
-
6. I w a m o t o M., N a g a s h i m a H., N a g a m i n e T. e.a. p-SINE-like intron of the CatA catalase homologs and phylogenetic relationships among AA-genome Oryza and related species. Theor. Appl. Genet., 1999, 98: 853-861.
-
7. T e o C.H., T a n S.H., H o C.L. e.a. Genome constitution and classification using retrotransposon-based markers in the Orphan Crop Banana. J. of Plant Biology, 2005, 48: 96-105.
-
8. K a l e n d a r R., G r o b T., R e g i n a M. e.a. IRAP and REMAP: two new retrotransposon-based DNA fingerprinting techniques. Theor. Appl. Genet., 1999, 98: 704-711.
-
9. Д р и б н о х о д о в а О.П., К о к а е в а З.Г., Г о с т и м с к и й С.А. Идентификация сортов, линий и мутантов гороха посевного с помощью RAPD-маркеров. С.-х. биол., 2005, 5: 61-66.
-
10. T o r r e s A.M., W e e d e n N.F., M a r t i n A. Linkage among sozyme, RFLP and RAPD markers in Vicia faba. Theor. Appl. Genet., 1993, 5: P.937-945.
-
11. P e a r c e S.R., S t u a r t - R o g e r s C., K n o x M.R. e.a. Rapid isolation of plant Ty1-copia group retrotransposon LTR sequences for molecular marker studies. The Plant Journal, 1999, 19: 711-717.
-
12. N e i M., L i W.-H. Mathematical model for studying genetic variation in terms of restriction endonucleases. Proc. Nat. Acad. Sci. USA, 1979, 76: 5269-5273.
-
13. V a n d e P e e r Y. TREECON for Windows: a software package for the construction and drawing of evolutionary trees for the Mi-
crosoft Windows environment. Computer Application in the Biosciences, 1994, 10, 5: 569-570.
-
14. F e l s e n s t e i n J. Confidence limits in phylogenies: an approach using bootstrap. Evolution, 1985, 39, 4: 783-791.
Московский государственный университет им. М.В. Ломоносова ,
