Особенности строения микробных образований в системе "риф - лагуна" (на примере верхнедевонского рифового комплекса Седъю, Южный Тиман)
")
Автор: Пономаренко Е.С.
Журнал: Вестник геонаук @vestnik-geo
Рубрика: Научные статьи
Статья в выпуске: 8 (296), 2019 года.
Бесплатный доступ
Описаны три типа микробных образований рифового комплекса Седъю, обнаженного на Южном Тимане: скелетные строматолиты, тонкозернистые строматолиты и микробные сгустковые известняки. Скелетные строматолиты построены оксигенными фототрофами - цианобактериями - и составляют значительную часть рифа Седъю. Сгустки являются результатом жизнедеятельности древних гетеротрофных бактерий и широко распространены в лагунных фациях. Тонкозернистые строматолиты занимают промежуточное положение. Выявлена зависимость распространения разных типов микробных образований от преобладания разных функциональных групп бактерий. Распространение бактерий, в свою очередь, регулировалось трофическими связями. Разработанная гипотетическая схема трофической структуры рифового комплекса Седъю показала наличие крупной трофической петли между рифом и лагуной, которая в общих чертах повторяет современную.
Система "риф - лагуна", микробные образования, трофические связи, южный тиман, франский ярус
Короткий адрес: https://sciup.org/149129393
IDR: 149129393 | УДК: 551.734.5[552.54+551.314] | DOI: 10.19110/2221-1381-2019-8-14-19
Features of microbial structures in the "reef - lagoon" system (on example of the Upper Devonian Sedyu reef complex, Southern Timan)
Three types of microbial structures of the Sedyu Reef Comples, exposed in the Southern Timan, were described: skeletal stromatolites, fine-grained stromatolites, and microbial clotted limestones. Skeletal stromatolites were constructed by oxygenic phototrops, cyanobacteria, and made up a significant part of the Sedyu Reef. Clots were resulted from vital activity of ancient heterotrophic bacteria and widespread in the lagoon facies. Fine-grained stromatolites occupied an intermediate position. The dependence of the distribution of different types of microbial structures on the predominance of different functional groups of bacteria was revealed. The bacteria spread, in turn, were regulated by trophic links. The developed hypothetical scheme of the trophic structure of the Sedyu Reef Complex showed the presence of a large trophic loop between the reef and the lagoon, which, in general terms, repeated the modern one.
Текст научной статьи Особенности строения микробных образований в системе "риф - лагуна" (на примере верхнедевонского рифового комплекса Седъю, Южный Тиман)
Девонские отложения Южного Тимана представлены разнофациальными отложениями. Интереснейшим геологическим объектом Южного Тимана является верх-нефранская толща на р. Седъю в районе одноименного поселка, интерпретируемая как ископаемый риф [8, 17 и др.]. В результате исследований было установлено, что большая часть верхнедевонских рифогенных карбонатов (в том числе и рифовый комплекс Седъю) сложена синезелеными водорослями (цианобактериями) [4, 12 др.] и сгустковыми известняками с фенестрами.
Роль микроорганизмов в осадочном процессе чрезвычайно велика. С микроорганизмами связаны многие месторождения полезных ископаемых, в частности джеспилиты, железомарганцевые образования, фосфориты и многие другие [1, 3, 7 и др.]. Огромное влияние бактериальные сообщества оказывают на карбонатное осадкообразование [16, 20, 22 и др.]. Объективная необходимость обобщения и систематизации для построения общей теории осадочного процесса с учетом бактериального фактора в настоящий момент образует новые направления — литологию микробиолитов [7] и бактериальный литогенез [1].
Цель настоящей работы состоит в установлении литологических особенностей разных типов известняковых микробных образований верхнедевонского рифового комплекса Седъю, закономерностей и вероятных причин их пространственного распределения.
Материал и методы
Верхнедевонский (верхнефранский) рифовый комплекс Седъю на р. Седъю в районе одноименного поселка вскрывается на протяжении нескольких километров в естественных выходах и карьерах (изучался автором летом 2011 г.). Здесь выделяются фациальные зоны (рис. 1) открытого моря, рифовых отмелей и лагун [9]. Микробные карбонаты в большом количестве отмечаются в рифовой и лагунной фациальных зонах.
Рифогенные образования изучались в обн. 12 и 14. Они представляют собой выходы в виде скал в верхней части склона и сложены вторичными пористыми и кавернозными доломитами с теневой структурой строматолитовых, биолитокластовых известняков, реже сгустковых узорчатых известняков. На нескольких уровнях развиты эрозионные поверхности [9].
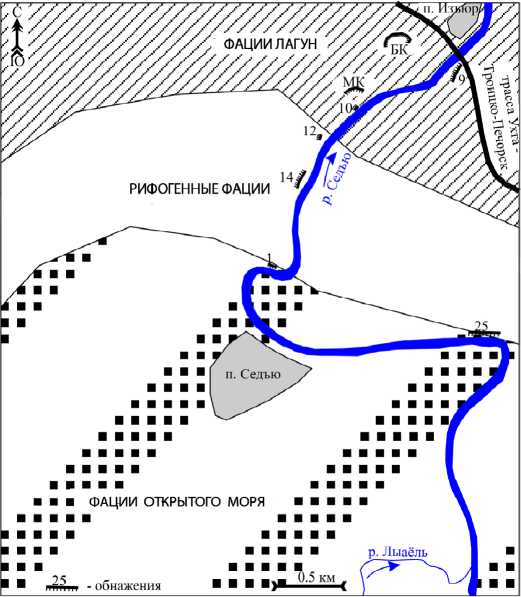
Рис. 1. Литолого-фациальный план-схема верхнефранских отложений в бассейне р. Седъю. МК — малый карьер, БК — большой карьер
Fig. 1. Lithologic-facial plane-scheme of Upper Fransian deposits in the Sedyu River basin. MK — small quarry, БК — big quarry
Лагунные отложения вскрыты северо-восточнее рифогенных (рис. 1). Ранее они интерпретировались как тыловая часть рифогенных фаций [8 и др.] или как образования полуизолированных лагун [2, 9]. В настоящей работе микробные известняки лагун рассматриваются на примере обн.10 и разрезов Малый Карьер и Большой Карьер (рис. 1). Лагунные отложения имеют пестрый литологический состав: известняки сгустковые, литокластовые, ооидные, пелитоморфные с редкими фаунистическими остатками (гастроподы, остракоды, амфипоры). Иногда отмечаются онкоиды.
Из рифогенных и лагунных образований было отобрано около 400 образцов; из них около 200 представлены микробными известняками или их доломитизированны-ми разностями. Изучение образцов производилось в шлифах и пришлифовках. Шлифы изучались под оптическим микроскопом ПОЛАМ РП-1 (ЛОМО), оснащенным цифровой камерой IS-500. Изученный материал хранится в геологическом музее им. А. А. Чернова в Институте геологии имени акад. Н. П. Юшкина (колл. № 663).
Типы микробных образований
Среди изученных микробных образований выделяются строматолиты и сгустки. Среди строматолитов по классификации Р. Райдинга (R. Riding) [30] можно выделить скелетные и тонкозернистые типы.
Скелетные строматолиты широко развиты в рифовой фации и сильно доломитизированы. Визуально они имеют кавернозно-пористое строение, слоистость подчеркивается чередованием светлых и темных слойков, хорошо заметных в пришлифовках (рис. 2, d). Форма строматолитов раз нообразная, чаще уплощенная, куполовидная. Наиболее крупные из них в центральной части массива достигают высоты до 0.1 м и длины до 0.5 м. В северной части рифовой зоны строматолиты более редки и не превышают 4— 5 см. Пространство между ними заполнено разнозернистым доломитом с теневыми структурами радиально-фибровых крустификационных корок (до 0.5—0.7 см), а также биокластов и литоклатов. В некоторых случаях среди строматолитов отмечаются колонии рецептакулит. Неизмененные биогермные породы были встречены в нижней по реке части обн. 25 и в развалах средней части склона обн. 12 (рис. 1), что позволило изучить микроструктуру самих строматолитов. В их строении также отмечено чередование темных и светлых слойков (рис. 2, а). Толщина темных слойков часто изменяется (от 0.06 до 0.4 мм). Они сложены плотным пелитоморфным кальцитом и распространены неравномерно: вверх по строматолиту их толщина увеличивается, в нижней части они часто выглядят как тонкие длинные линзы. Толщина светлых слойков, наоборот, от основания к верхней части строматолита уменьшается от 1.1 мм до 0.06 мм. Светлые слои сложены разнозернистым спари-товым кальцитом (от 0.007 до 1.0 мм), более крупные кристаллы ориентированы по наслоению. По распределению зерен кальцита в них часто можно выделить более тонкие слойки (0.01—0.02 мм толщиной). Кроме того, в светлых слойках отмечаются трубчатые образования (рис. 2, с), по форме напоминающие нитчатые цианобактерии. На строматолитах нарастают колонии цианобактерий Schuguria flabelliformis Antrop. (рис. 2, b), значительно реже — небольшие (до 0.3 мм) кустистые колонии, напоминающие Epiphyton.
Интерпретация. Характер строения изученных неизмененных строматолитов, в которых угадываются реликты нитчатых цианобактерий, позволяет относить их к типу скелетных строматолитов [22]. Таким образом, формирование биогермных пород происходило за счет жизнедеятельности бактериальных сообществ, где ведущая роль была отведена оксигенным фототрофам — цианобактериям. Ведущая роль цианобактерий как первичных продуцентов признается для многих строматолитов.
Тонкозернистые строматолиты занимают промежуточное положение (обн. 10) между скелетными строматолитами и сгустковыми известняками лагунных фаций. Они имеют столбчатую или куполовидную формы (рис. 2, h) и в основном представлены чередованием темных слойков до 0.2 мм толщиной, сложенных плотным пятнистым пелитоморфным кальцитом, внутри которых отмечаются сильно измененные кальцисферы и теневые нитчатые образования или колонии цианобактерий (до 1.0—1.5 см) Garwoodia? (рис. 2, i). Между строматолитами развит литокластовый материал, связанный спаритовым кальцитовым цементом.
Интерпретация. Согласно классификации [22], эти строматолиты относятся к типу тонкозернистых. В строении современных тонкозернистых строматолитов принимают участие различные функциональные группы бактерий [3, 20 и др.]. Тонкозернистые строматолиты в рифовом комплексе Седъю развиты между скелетными строматолитами и сгустковыми известняками и, следовательно, представляют собой переходный тип микробиальных образований. В их строении отмечаются тени колоний и нитей цианобактерий, характерных для скелетных строматолитов, также большую роль играет пелитоморфный кальцит, слагающий сгустки в лагунных фациях.
Сгустки наиболее широко распространены в разрезах лагунной фации и слагают сгустковые известняки. Макро- скопически они имеют плитчатое строение (плитки до 0.5 м), цвет пород от светло-кремово-серых до серых. Иногда отмечается тонкая неправильная слоистость (рис. 2, е), вследствие чего породы сходны с ламинитами — микробными образованиями литорали. Микроскопически эти породы сложены мелкими (0.1—0.2 мм) пелоидами с нечеткими облаковидными краями и кальцисферами, между которыми в порах развит спаритовый кальцит. Такая раз новидность пелоидов или сгустков в настоящее время многими исследователями относится к продуктам жизнедеятельности бактериальных сообществ [2, 21—23 и др.]. Часто сгустки образуют агрегаты более крупного размера разных форм (кустистые и древовидные), реже более крупные ветвящиеся столбчатые разности (рис. 2, f). Исследование сгустков методом сканирующей электронной микроскопии [10] показало, что они сложены мелкими кристаллами каль-
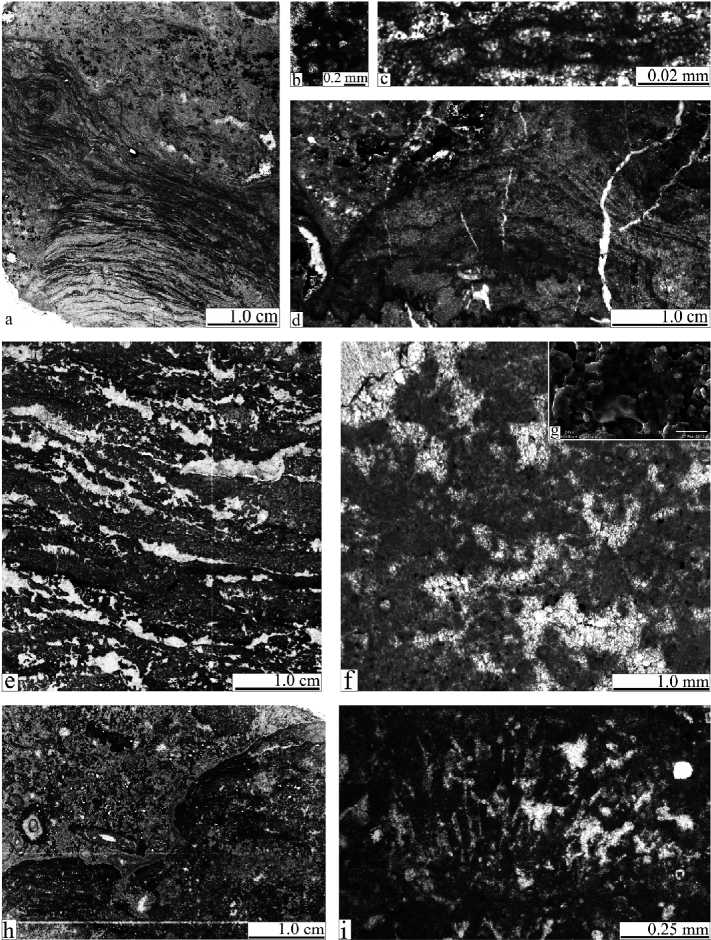
Рис. 2. Основные типы литифицированных микробных образований: а — строматолитовый известняк с разнообразными остатками цианобактерий. Шлиф С12/18с-2011; b — колония пузырчатой цианобактерии Schuguria . Шлиф С12/18с-2011; с — теневые нитчатые образования в строматолите. Шлиф С12/18с-2011; d — вторичный доломит с теневой структурой строматолита. Обр. С14/19-2011. Пришлифовка; е — фенестровый сгустковый микробный известняк с тонкой неправильной слоистостью. Шлиф С4/74-2011; f — кустовидные агрегаты из микробных сгустков. Шлиф С4/97-2011; g — строение сгустков под сканирующим электронным микроскопом. Отмечаются многочисленные мелкие зерна кальцита, покрытые микробной пленкой. Обр. С4/74-2011; h — тонкозернистый строматолит в онкоидно-литокластовом известняке. Шлиф С11 /1а-2011; i — теневая структура кустистой колонии цианобактерии (вероятно, Garwoodia ) в тонкозернистом строматолите. Шлиф МК7б-2011
Fig. 2. Main types of lithified microbial structures: a — stromatolite limestone with various cyanobacteria fossils. Thin-section C12/18c-2011; b — colony ofthe vesicular cyanobacreia Schuguria . Thin-section C12/18c-2011; c — shadow filamentous structures in the stromatolite. Thin-section C12/18c-2011; d — secondary dolomite with shadow of the stromatolite. Sample C14/19-2011. Polished section; e — fenestral clotted microbial limestone with thin irregular lamination. Thin-section C4/74-2011; f — bush-like aggregates from microbial limestones. Thin-section C4/97-2011; g — a clots structure under SEM. There are numerous small calcite grains coated with a microbial film. Sample C4/74-2011; h — fine-grained stromatolite in the oncoid-lithoclastic limestone. Thin-section C11 /1a-2011; i — a shadow structure of a bushy colony of cuanobacteria (probably, Garwoodia ) in the fine-grained stromatolite. Thin-section МК7и-2011
пита (до 2.0 pm), связанными пленками (рис. 2, g), по всей видимости бактериального генезиса, из-за чего кристаллический кальцит имеет сглаженные очертания.
Интерпретация . Ранее сгустки рассматривались как кальцифицированные ЭПС (Extracellular Polymeric Substances) [22], но в настоящий момент [23] эти зерна интерпретируются как продукт гетеротрофной бактериальной кальцификации ЭПС и других клеточных продуктов. Полевыми и лабораторными работами [20] установлено, что самая верхняя часть нитей цианобактерий вблизи к поверхности бактериального мата обычно не обызвествлена, а кальцитизация возрастает книзу по направлению к зоне разложения. К похожим выводам пришли и другие исследователи [5, 14, 19], указав, что образование известковой оболочки происходит в плохо функционирующих или отмерших частях мата. Широкие масштабы бактериальных процессов приводят к концентрации НСО3- и увеличению щелочности, что благоприятствует осаждению СаСО3 [24, 30]. Это связано с процессами органоминерализации — образования минералов при влиянии органического вещества вне зависимости от живущих организмов, из которых они были получены [15]. Тафономические изменения цианобактерий под воздействием гетеротрофной бактериальной активности были показаны в работах различных исследователей [13, 21, 24 и др.]. По предложенной модели [21], свободные нити цианобактерий ( Girvanella ) кальцифицируются, а затем под действием разложения превращаются в сгустки (рис. 3, а).
Приведенные выше данные свидетельствуют об образовании сгустковых известняков, которые широко развиты, в том числе и в лагунных фациях рифового комплекса Седью, при значительном участии гетеротрофных бактерий, разлагавших, по всей видимости, слизь цианобактерий. В пользу последнего предположения свидетельствует кустис
Кальцификация
Разложение
Разложение
Свободные нити
Кальцификация
Girvanella
РИФ
Ц_И АНОБАКТЕР1Л
Кальцификация Седиментация
нити Girvanella
ЛАГУНА
РЕЦЕПТАКУЛИТЫД
тая и древовидная форма некоторых сгустковых агрегатов, напоминающая колонии Schuguria и Epiphyton (рис. 2, f).
Таким образом, анализ распространения основных типов микробных известняков ясно указывает на наличие скелетных строматолитов, строивших основную часть верхнефранского рифа Седью. В лагунных отложениях остатки бактериальных сообществ уже представлены в основном сгустками. Эти типы микробных образований, по всей видимости, зависят от жизнедеятельности функционально различных групп бактерий: в скелетных строматолитах ведущую роль играли оксигенные фототроф-ные цианобактерии, тогда как сгустки являются результатом жизнедеятельности гетеротрофных бактерий. Тонкозернистые строматолиты занимают промежуточное положение, как по своему строению, так и по положению (на границе рифа и лагуны).
Связь трофической структуры в системе «риф — лагуна» и типами микробных образований
Для современной рифовой системы [11] характерно наличие двух важнейших, пространственно разнесенных, но связанных функционально трофодинамических ступеней — продуцирующей (сложенной главным образом автотрофными организмами) и потребляющей (образуемой комплексом гетеротрофных организмов, которые утилизируют биомассу, создаваемую первой трофической ступенью). Растворенный и взвешенный органический материал, поступающий из автотрофных зон, волнами забрасывается в лагуну, где заселяется бактериальными сообществами и осаждается на дно. Этими агрегатами питаются многочисленные планктонофаги, что приводит к парадоксу обилия планктонофагов в рифовых лагунах при крайней бедности планктона [6, 11].
На примере верхнефранского рифового комплекса Седью автотрофная зона представлена скелетными строматолитами. Преобладание оксиген-ных фототрофов (цианобактерий) свидетельствует о том, что сообщество этой зоны обладало собственной первичной продукцией. О наличии гетеротрофной зоны в лагуне свидетельствует обилие сгустков — результата жизнедеятельности гетеротрофных бактерий. Ана-
Тромбоиды
ГЕТЕРОТРОФНЫЕ БАКТЕРИИ
Рис. 3. а — тафономические изменения микробных образований (по данным Pratt, 1995, fig. 62); b — типы микробных образований в верхнефранс-ких карбонатных породах. Слева направо : колония цианобактерий в ассоциации со скелетными строматолитами, тень колонии цианобактерий в тонкозернистом строматолите, микробные сгустки; с — гипотетическая схема трофической структуры бентосных сообществ рифового комплекса Седью
Fig. 3. а — taphonomic changes of microbial structures (the Pratt model. 1995, fig. 62). b — types of microbial structures in the Upper Changes Frasnian carbonate rocks. From left to right : cyanobacterial colony in association with skeletal stromatolites, a shadow cyanobacteria colony in fine-grained stromatolite, microbial clots; с — a hypothetical scheme of the trophic structure of the benthic communities of the Sedyu Reef Complex
лиз специфики органических остатков и литологического состава лагунных фенестровых известняков позволили создать идеализированную схему трофических цепей в лагуне [2]. Гипотетическая схема трофической структуры и трофических цепей рифового комплекса Седъю, представленная на рис. 3, с, показывает наличие автотрофно/гетерот-рофной трофической петли между рифом и лагуной. Органика, произведенная автотрофными цианобактериями, использовалась гетеротрофными бактериями в лагуне, полученный при разложении органики СО2 мог быть использован оксигенными фототрофами на рифе. Внутри лагуны выделяется несколько гетеротрофных трофических петель, что характерно для детритных пищевых цепей.
Таким образом, распределение разных типов микробных известняков зависело от преобладающего типа бактериальных сообществ, чьё распространение, в свою очередь, регулировалось трофическими связями.
Заключение
Таким образом, результаты исследований и их интерпретацию можно кратко изложить в следующих пунктах:
-
1. В изученном верхнефранском рифовом комплексе Седъю большую роль играют микробные образования. Их распространение имеет явную фациальную зависимость: скелетные строматолиты являются основными строителями рифа, на границе с лагуной отмечаются тонкозернистые строматолиты, а в лагуне развиты микробные сгуст-ковые известняки.
-
2. В скелетных строматолитах отмечаются тени нитчатых цианобактерий, а они сами ассоциируются с ясно заметными колониями пузырчатых и кустистых цианобактерий. Анализ опубликованной литературы показал, что за образование сгустковых микробных известняков ответственны гетеротрофные бактериальные сообщества. Согласно модели Пратта (Pratt, 1995), сгустки представляют собой результат тафономических изменений цианобактерий под воздействием гетеротрофной бактериальной активности. Это подтверждается фактическим материалом: в шлифах некоторые агрегаты из сгустков по форме напоминают пузырчатые и кустистые колонии цианобактерий.
-
3. Присутствие скелетных строматолитов и многочисленных других цианобактерий в рифогенных породах свидетельствует о наличии продуктивной автотрофной зоны на рифе, тогда как в лагуне преобладала гетеротрофная составляющая палеоэкосистемы. Это указывает на наличие трофической петли между позднедевонским строматолитовым рифом и его лагуной и в общих чертах повторяет структуру внешней трофической петли современных рифов. Пространственное распределение разных типов микробных известняков, таким образом, зависело от преобладающего типа бактериальных сообществ, чьё распространение, в свою очередь, регулировалось трофическими связями.
Работа проводилась в рамках Госпрограммы № АААА-А17-117121270034-3 и при частичной финансовой поддержке Программы фундаментальных исследований УрО РАН, проект №18-5-5-31.
Список литературы Особенности строения микробных образований в системе "риф - лагуна" (на примере верхнедевонского рифового комплекса Седъю, Южный Тиман)
- Антошкина А. И. Бактериальный литогенез // Обзор концептуальных проблем литологии. М.: ГЕОС, 2012. С. 89-105.
- Антошкина А. И. Пономаренко Е. С., Канева Н. А. Фенестровые известняки как индикатор специфики позднедевонских морей, Тимано-Североуральский регион // Литология и полезные ископаемые. 2014. №. 6. С. 493-505.
- Бактериальная палеонтология / Под ред. А. Ю. Розанова. М.: ПИН РАН, 2002. 188 с.
- Беляева Н. В., Корзун А. Л., Петрова Л. В. Модель седиментации франско-турнейских отложений на северо-востоке Европейской платформы (в связи с формированием рифовых резервуаров). СПб.: Наука, 1998. 154 с.
- Герасименко Л. М. Фотосинтез и актуалистическая палеонтология цианобактерий // Проблемы ранней эволюции фотосинтеза. М.: ПИН РАН, 2011. С. 37-65. (Геобиологические процессы в прошлом).
