Особенности захоронения младенцев в жилищах городища Чича-1 в Барабинской лесостепи по данным анализа структуры ДНК

Автор: Пилипенко А.С., Ромащенко А.Г., Молодин В.И., Куликов И.В., Кобзев В.Ф., Поздняков Д.В., Новикова О.И.
Журнал: Археология, этнография и антропология Евразии @journal-aeae-ru
Рубрика: Эпоха палеометалла
Статья в выпуске: 2 (34), 2008 года.
Бесплатный доступ
В IX-VIIee. до н.э., в переходный период от бронзового к железному веку, на территории Барабы возникает крупное городище Чича-1. Новые черты социальной организации населения городища проявились и в практике захоронения младенцев непосредственно в жилищах. В настоящей работе предпринята попытка методами анализа структуры ДНК выяснить мотивации отбора младенцев. Результаты анализа половой принадлежности погребенных указывают на осознанный выбор мальчиков для захоронения в жилищах. Наличие разных вариантов гаплотипов митохондриальнои ДНК у двух младенцев, захороненных в одном жилище, свидетельствует об отсутствии между ними прямого родства по материнской линии. Некоторые из обнаруженных гаплотипов мтДНК младенцев, нехарактерные для древнего и современного населения региона, в настоящее время распространены на территориях, простирающихся к югу и юго-западу от Барабы (Средняя и Передняя Азия, Ближний Восток, Кавказ).
Короткий адрес: https://sciup.org/14522653
IDR: 14522653 | УДК: 902
Текст научной статьи Особенности захоронения младенцев в жилищах городища Чича-1 в Барабинской лесостепи по данным анализа структуры ДНК
В IX–VII вв., в переходный период от бронзового к железному веку, в лесостепной зоне Западной Сибири происходили процессы, в определенной степени сопоставимые с цивилизационными [Молодин и др., 2004]. В это время в Барабинской лесостепи появилось население, мигрировавшее, по-видимому, с запада, юго-запада. В результате здесь сформировалась смешанная в этнокультурном плане популяция с новыми (привнесенными) чертами социальной организации.
В предшествовавшие эпохи развитой и поздней бронзы на территорию Барабы уже проникало население из юго-западных районов современных Средней Азии и Казахстана. Наиболее ранний импульс миграции относится к самому началу эпохи развитой бронзы, по-видимому, к рубежу III–II тыс. до н.э., о чем свидетельствуют обнаруженные на могильнике Сопка-2 пастовые бусы-крестовики, сердоликовые бусины со следами травления, а также другие предметы из Средней Азии. Антропологи отмечают появление специфических европеоидных черт в костном материале [Молодин, 1988]. Следующая волна миграции относится ко второй половине II тыс. до н.э.: автохтонное население – представители кротовской культуры – сменилось пришлыми европеоидными носителями андроновской (федоровской) культуры.
В результате длительного сосуществования носителей пришлой андроновской и аборигенной культур сформировался пласт населения, представлявшего ир-менскую культуру эпохи поздней бронзы [Молодин, 1985], которая, по сути, стала автохтонной для региона. В конце эпохи бронзы в Барабинскую лесостепь и Кулундинскую степь устремилось несколько мигра-
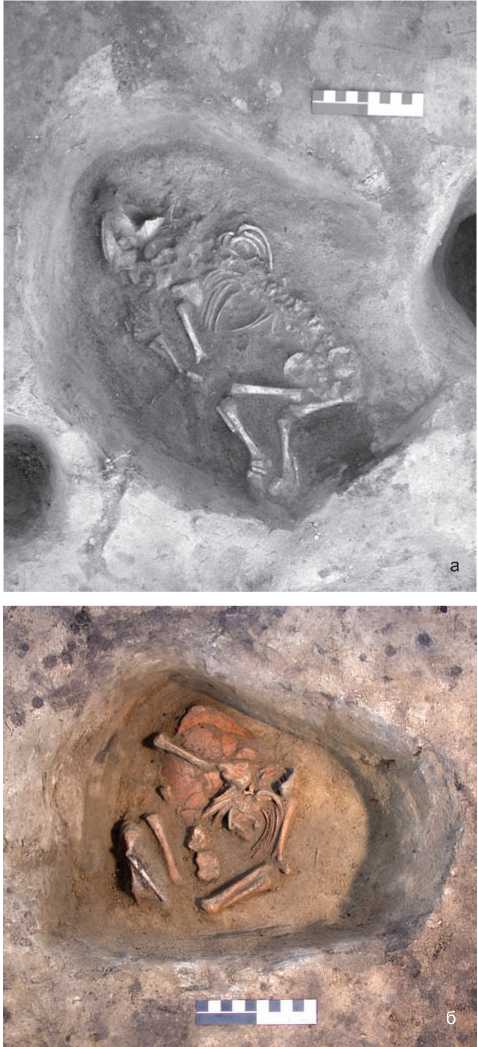
Рис. 1. Городище Чича-1, раскоп 10, жилище 10, погребение 1 (а) и 2 (б). Вид с юга.
ционных потоков с запада и юго-запада. Сначала это были носители культуры валиковой керамики [Кирюшин, Удодов, 1992, с. 89], а затем бегазы-дандыбаев-ской [Молодин, 1981, с. 15–17; Удодов, 1988, с. 107–110]. Следы еще одного потока мигрантов с юго-запада, относящегося к переходу от бронзового к железному веку, зафиксированы на городище Чича-1. Здесь пришлое население сосуществовало с автохтонным позднеирмен- ским, вступая с последним, по-видимому, не только в культурные, но и в кровно-род-ственные отношения [Молодин, 2006б]. Отмеченные выше миграционные потоки, двигавшиеся в эпоху бронзы через Барабин-скую лесостепь, несомненно, стали причиной чрезвычайно сложной этнокультурной ситуации в регионе.
Постепенно у носителей позднеирменской культуры сформировались новые формы социально-культурной организации. Они проявились, например, в создании крупного городища Чича-1, элементах до-мо строительства, возведении мощных фортификационных сооружений, зачатках ремесленничества и, по всей видимости, в изменении некоторых сторон духовной культуры [Молодин и др., 2004]. Новации нашли отражение в погребальной практике. Удалось выявить некрополи, оставленные как автохтонным населением, так и мигрантами [Молодин, 2006а].
В этой связи особый интерес вызывают захоронения младенцев, обнаруженные на памятнике Чича-1 в отдельных жилых домах как аборигенного, так и пришлого населения (рис. 1). Молекулярно-генетические исследования структуры ДНК этих погребенных должны помочь определить истоки пришлого населения городища Чича-1, т.к. подобный обряд не был характерен для коренных жителей Барабы предшествовавших исторических периодов. Он был широко распространен у представителей древнеземледельческих культур Средней и Передней Азии [Алекшин, 1986, с. 151–153]. В литературе этот феномен связывают со строительными жертвоприношениями [Байбу-рин, 1983, с. 61–62; Формозов, 1984, с. 240; Кузьмина, 1994, с. 97], с обрядами, направленными на предотвращение болезни и смерти будущих детей [Антонова, 1990, с. 105], идеей возвращения умершего [Там же; Алекшин, 1986, с. 152], культом плодородия [Бибиков, 1953, с. 197–198; Антонова, 1990, с. 106–107]. Имеются этнографические источники, свидетельствующие о захоронении детей по особому обряду, в основе которого лежат представления о принадлежности ребенка, не достигшего определенного возраста, еще не к миру людей, а к миру духов, в связи с чем он наделялся некими мистическими качествами. Так, у кар-гинцев детей заворачивали в кошму, затем в бересту и подвешивали на дереве или клали в дупло [Усманов, 1980, с. 109]. Воздушное захоронение на дереве мертворожденных или умерших младенцев устраивали барабинские татары [Титова, 1976, с. 132]. У обских угров умерших детей, души которых считались очень опасными [Карьялайнен, 1994], хоронили на отдельных кладбищах под корнями и в дуплах деревьев завернутыми в бересту [Соколова, 1980, с. 130, 137]. Вместе с тем в этнографии народов Сибири отсутствуют сведения о погребении умерших детей в жилых домах. Очевидно, данная традиция здесь не имела места. В настоящей работе предпринимается попытка изучить особенности зафиксированного на городище обряда захоронения младенцев методами молекулярно-генетического анализа структуры их ДНК.
Материалы и методы
Для выделения ДНК использовались фрагменты скелетов восьми из десяти младенцев, захоронения которых были обнаружены в жилищах городища Чича-1 [Молодин и др., 2004] (табл. 1). В жилищах находилось по одному или два погребения. В выборке представлены образцы костей младенцев из разных частей городища: четырех из «цитадели» (площадка II) (рис. 2) – зоны проживания автохтонного для того периода населения; четырех с периферии (один погребенный с площадки III и трое с площадки IV), заселенной, вероятно, мигрантами [Там же]. Захоронения двух детей в одном жилище представлены парами образцов – № 3, 4 (жилище № 20, площадка II) и 6, 7 (жилище № 10, площадка IV).
Для исследования от шести из восьми скелетов использовались целые бедренные ко сти хорошей макроскопической сохранности, от двух других скелетов – фрагменты трубчатых костей более низкой степени сохранности.
Предварительная обработка костного материала и выделение ДНК. Поверхность костей механи-че ски очищали от загрязнений и чужеродной ДНК. После удаления внешнего слоя костной ткани (ок. 1– 2 мм) поверхность обрабатывали перекисью водорода или гипохлоритом натрия и облучали ультрафиолетовым светом (30 мин с каждой стороны). Из внутренней области костных фрагментов высверливали мелкодисперсный костный порошок, из которого экстрагировали ДНК.
Тотальную ДНК выделяли обработкой костного порошка 5М гуанидинизотиоционатным буфером pH 11 (48 ч при температуре 65 оС) и последователь- ной экстракцией сначала смесью фенола и хлороформа, а затем хлороформом. Осаждение ДНК из водной фазы в присутствии 1М NaCl осуществляли изопропиловым спиртом. ДНК хранили в водном растворе при температуре –20 оС.
Определение половой принадлежности погребенных. Пол определяли путем амплификации с помощью полимеразной цепной реакции (ПЦР) фрагментов ДНК, специфичных для хромосом X и Y перицентромерных альфоидных повторяющихся последовательностей нуклеотидов, размером 130 пар нуклеотидов (п.н.) (из последовательности DXZ1 размером 2,0 тыс. п.н. ~5 тыс. копий на хромосоме Х) и 170 п.н. (из последовательности DYZ3 размером 5,5 тыс. п.н. ~1 тыс. копий на хромосоме Y). ПЦР, специфичные для фрагментов ДНК Х- и Y-хромосом, проводили в разных пробирках. Для амплификации использовали соответствующие пары праймеров: X1 (5`-aatcatcaaatggagatttg-3`), X2 (5`-gttcagctctgtgagtgaaa-3`) для Х-хромосомы, Y1 (5`-atgatagaaacggaaatatg-3`), Y2 (5`-agtagaatgcaaagggctc-3`). Реакционная смесь объемом 25 мкл включала: 75 мМ Tris HCl (pH 9,0); 20 мМ (NH4)2SO4; 0,01 % Tween-20; 3,0 мМ MgCl2; 0,2 мМ каждого dNTP; 1 μM каждого праймера; 1 мг/мл БСА; 5 мкл раствора тотальной ДНК; 0,75 единиц активности (ед. акт.) термостабильной ДНК-полимеразы. Режим амплификации: 95 оС 3 мин и 42 цикла – 95 оС 30 с, 55 оС 30 с, 72 оС 30 с. Результаты реакции визуализировали разделением продуктов амплификации в 4%-м полиакриламидном геле (ПААГ) с последующим окрашиванием их бромистым этидием. При облучении геля УФ наблюдали специфические полосы: 130 п.н. – для Х-хромосомы и 170 п.н. – для Y-хромосомы. Если в образце ДНК обнаруживали специфические последовательности как Х-, так и Y-хромосомы, – пол индивида мужской; если обнаруживали ДНК Х-хромо сомы и отсутствие продукта для Y-хромосомы (только полоса 130 п.н.), – пол женский.
Таблица 1. Палеоантропологический материал с городища Чича-1, использовавшийся для анализа ДНК
|
Номер образца |
Место захоронения останков |
Фрагмент скелета |
|
1 |
Площадка II, раскоп 6, жилище 3а, погр. 2 |
Бедренная кость |
|
2 |
» раскоп 7, жилище 9, погр. 1 |
» » |
|
3 |
» раскоп 17, жилище 20, погр. 1 |
» » |
|
4 |
» раскоп 17, жилище 20, погр. 2 |
» » |
|
5 |
Площадка IIIb, раскоп 5, жилище 8, погр. 1 |
Фрагменты трубчатых костей |
|
6 |
Площадка IV, раскоп 10, жилище 10, погр. 1 |
Бедренная кость |
|
7 |
То же, погр. 2 |
» » |
|
8 |
Площадка IV, раскоп 12, жилище 11, погр. 2 |
Фрагменты трубчатых костей |
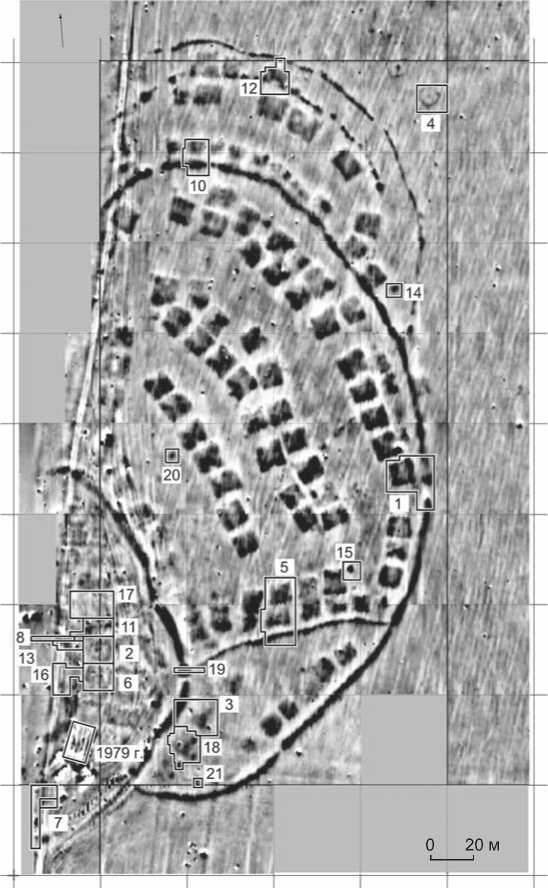
Рис. 2. Магнитограмма городища Чича-1 с выделенными границами раскопов (цифры соответствуют их номерам).
Амплификация ГВС I митохондриальной ДНК. Амплификацию последовательности нуклеотидов первого гипервариабельного сегмента некодирующей области мтДНК (ГВС I мтДНК) проводили, используя два варианта ПЦР. Метод «вложенной ПЦР» (nested PCR) включает в себя два последовательных раунда ПЦР. Первый раунд ПЦР с внешними праймерами L16046 (5`-ttctttcatggggaagcagattt-3`) и H16401 (5`-attgatttcacggaggatggtg-3`) проводили в объеме 25 мкл. Реакционная смесь включала: 75 мМ Tris HCl (pH 9,0); 20 мМ (NH4)2SO4; 0,01 % Tween-20; 5,0 мМ MgCl2; 0,2 мМ каждого dNTP; 1 μM каждого праймера; 1 мг/мл БСА; 5 мкл раствора тотальной ДНК; 0,75 ед. акт. термостабильной ДНК-полимеразы. Реакцию проводили в следующем режиме: первоначальная денатурация – 95 оС 3 мин, затем 10 циклов – 95 оС 1 мин, 55 оС 1 мин, 72 оС 1 мин и 30 циклов в режиме – 95 оС 23 с, 55 оС 23 с, 72 оС 23 с. Второй раунд ПЦР (продукт 336 п.н.) проводили в объеме 50 мкл с внутренними праймерами L16073 (5`-ccacccaagtattgactcaccc-3`) и H16367 (5`-ctatctgaggggggtcatccat-3`). Реакционная смесь включала: 75 мМ Tris HCl (pH 9,0); 20 мМ (NH4)2SO4; 0,01% Tween-20; 5,0 мМ MgCl2; 0,2 мМ каждого dNTP; 1 μM каждого праймера; 4 мкл продукта первого раунда ПЦР и 1,5 ед. акт. термостабильной ДНК-полимеразы. Реакцию проводили в следующем режиме: 95 оС 3 мин и 25 циклов – 95 оС 23 с, 55 оС 23 с, 72 оС 23 с.
Амплификацию 5`-участка ГВС I (между позициями 16004–16193 кембриджской референсной последовательности) проводили с праймерами L16004 (5`-ccattagcacccaaagcta agattc-3`) и H16193 (5`-gtacttgcttgtaagcatg-3`) [Adcock, Dennis, Easteal, 2001]. Реакционная смесь объемом 50 мкл включала: 75 мМ Tris HCl (pH 9,0); 20 мМ (NH4)2SO4; 0,01 % Tween-20; 5,0 мМ MgCl2; 0,2 мМ каждого dNTP; 1 μM каждого праймера; 1 мг/мл БСА; 5 мкл раствора тотальной ДНК; 1,5 ед. акт. термостабильной ДНК-полимеразы. Режим амплификации: 95 оС 3 мин и 40 циклов – 95 оС 30 с, 55 оС 30 с, 72 оС 30 с.
Амплификацию участка второго гипервариабельного сегмента некодирующей области мтДНК (ГВС II) в сегменте 16517– 00160 проводили в реакционной смеси объемом 50 мкл, по составу аналогичной предыдущей, с праймерами L16517 (5`-catctggttcctacttcagg-3`) и H00160 (5`-tgtaatattgaacgtaggtgcgat-3`), в ре- жиме – 95 оС 3 мин и 42 цикла – 95 оС 30 с, 55 оС 30 с, 72 оС 30 с.
Детекцию продуктов ПЦР осуществляли электрофоретическим их разделением в 4%-м ПААГ с последующим окрашиванием бромистым этидием и облучением ультрафиолетовым светом.
Определение нуклеотидной последовательности амплифицированных участков мтДНК проводили прямым автоматическим секвенированием с использованием BigDye Terminator v3.1 Cycle Sequencing (Applied Biosystems, США), согласно рекомендациям производителя. Для секвенирования продукта ПЦР использовали секвенирующие праймеры L16114 (5`-ggggacgagaagggatttga-3`) и H16347 (5`-tttcgtacattactgccagccac-3`). Образцы ДНК анализировали на автоматическом секвенаторе ABI Prism 310 Genetic Analyzer (Applied Biosystems, США).
Полученные последовательности мтДНК из останков древних индивидов сравнивали с кембридж- ской референсной последовательностью мтДНК для выявления специфических нуклеотидных замен и определения гаплотипов [Anderson et al., 1981] с помощью пакета программ DNAStar (DNAStar Inc., США) и программы Sequence Scanner (Applied Biosistems, США).
Меры против контаминации . Все стадии работы с древним материалом проводили в специально оборудованном изолированном помещении с использованием специальной одежды, лицевых масок, очков, стерильных перчаток. Все рабочие поверх-но сти в помещении регулярно обрабатывали 5%-м раствором гипохлорита натрия и облучали ультрафиолетом. На всех этапах использовали стерильные реактивы и пластиковую посуду, наконечники для автоматических дозаторов с фильтрами. Экстракцию ДНК и постановку ПЦР контролировали на отсутствие загрязнений в виде чужеродной ДНК во всех компонентах растворов. Повторные экстракции ДНК из одних и тех же ко стных образцов осу-ще ствляли в разное время. Для всех работающих с древней ДНК сотрудников была определена последовательность нуклеотидов гипервариабельных сегментов I и II мтДНК.
Результаты
Пол младенцев из погребений в жилищах городища Чича-1. Во всех восьми образцах тотальной ДНК были выявлены специфические последовательности нуклеотидов как Х-, так и Y-хромосом. Пол каждого младенца определяли с использованием двух и более независимо выделенных образцов тотальной ДНК. ПЦР для каждого образца ДНК повторяли дважды. Во всех случаях контроли экстракции и чистоты ПЦР-системы указывали на отсутствие контаминации. Таким образом, нами достоверно установлен мужской пол всех младенцев, подвергнутых исследованию (табл. 2).
Гаплотипирование мтДНК погребенных. Последовательности нуклеотидов ГВС I мтДНК были определены для шести образцов. Для образцов № 1, 2, 4, 6 дополнительно установлены последовательно сти нуклеотидов участка ГВС II в сегменте 16517 – 00160 и показано наличие в них транзиции A00073G. Сопоставлением с референсной последовательностью нуклеотидов контрольного района мтДНК были выявлены гаплотипы и определена их принадлежность к гаплогруппам, согласно общепринятой в настоящее время классификации [Richards et al., 1998, 2000] (табл. 2).
Гаплотипы мтДНК всех детей различаются. Следовательно, индивиды № 3, 4 и 6, 7, погребенные парами в жилищах № 20 и 10 соответственно, не могут являться прямыми родственниками по материнской линии. Данное обстоятельство позволяет предположить, что либо в некоторых жилищах Чичи-1 проживало несколько семей, либо у данного сообщества бытовала полигамная структура организации семьи.
Мужской пол всех исследованных младенцев и отсутствие родства по материнской линии у погребенных попарно указывают на возможность существования у населения городища Чича-1 специфического обряда, связанного с особым отношением к детям мужского пола. Последнее было присуще сообществам со скотоводческим укладом хозяйства и группам с присваивающей системой экономики и бытовало до этнографической современности [Литвинский, 1958, с. 31–33; Заднепровский, 1962, с. 99].
Принадлежность установленных гаплотипов мтДНК младенцев к разным западно-евразийским гаплогруппам свидетельствует об отсутствии существенного восточно-евразийского влияния на генофонд населения городища, а также о высокой степени их филогенетической неоднородности.
Таблица 2. Гаплотипы митохондриальной ДНК и половая принадлежность останков младенцев с городища Чича-1
|
Номер образца |
Гаплотип ГВС I |
Гаплогруппа мтДНК |
Число независимых экстракций ДНК |
Число секвенированных последовательностей ГВС I мтДНК |
Нуклеотид в положении 00073 ГВС II мтДНК |
Пол |
|
1 |
16093-16224-16311 |
K |
2 |
2 |
G |
Мужской |
|
2 |
16183A→C-16189-16249 |
U1a |
3 |
4 |
G |
» |
|
3 |
16189-16260-16270 |
U5b |
2 |
2 |
– |
» |
|
4 |
16356-16362 |
U4 |
3 |
6 |
G |
» |
|
5 |
– |
– |
– |
– |
– |
» |
|
6 |
16069-16126 |
J |
2 |
3 |
G |
» |
|
7 |
16366 |
H |
2 |
3 |
– |
» |
|
8 |
– |
– |
– |
– |
– |
» |
Обсуждение результатов
Городище Чича-1 является одним из наиболее крупных из известных за Уралом поселений переходного от бронзы к железу времени [Молодин и др., 2006]. Анализ артефактов, прежде всего керамики, выявил этнокультурную неоднородность его населения. Территория городища подразделяется на две основные зоны обитания – «цитадель» (площадки I, II) и периферию (площадки III, IV), соответствующие, вероятно, зонам проживания двух различных этнокультурных групп [Молодин, 2006б]. С «цитаделью» связан первый этап освоения территории городища автохтонным для Барабы населением (сначала ир-менцами, а затем и позднеирменцами) при участии мигрантов из различных регионов Западной Сибири [Молодин и др., 2004]. Освоение зон III и IV происходило при доминировании пришлого населения из западных и юго-западных районов, скорее всего, с территории современного Казахстана. По-видимо-му, длительное время две этнокультурные группы сосуществовали на поселении, сохраняя свою специфику культурно-хозяйственного уклада [Молодин, 2006б]. На городище выявлены также следы пребывания носителей культур с территорий, расположенных к северу и северо-западу от Барабы (атлымской, красноозерской, завьяловской и сузгунской). Это свидетельствует о том, что на поселении проживали также представители культур таежного и лесостепного поясов Западной Сибири. Весьма высока вероятность вступления носителей этих культурных образований в брачные контакты с основными обитателями городища, что делало этнокультурную ситуацию на памятнике еще более сложной.
Погребения, обнаруженные в жилищах, либо примыкали к восточной или западной стене котлована, либо находились вблизи от одного из углов жилища и, как правило, были вписаны в ряд между столбовыми ямами. То, что захоронение младенцев было произведено в жилищах намеренно, позволяет выдвинуть предположение о существовании у населения Чичи-1 специфического погребального обряда [Молодин и др., 2004]. Бесспорно, рассматриваемый обряд не распространялся на всех младенцев, умиравших на поселении. Для древнего населения конца эпохи бронзы был характерен высокий уровень детской смертности [Чикишева, 2000]. Расчеты, произведенные для могильника ирменской культуры Журавлево-4, показали, что 29 % всех погребенных составляли дети и подростки, причем преобладали младенцы [Бобров, Чикишева, Михайлов, 1993].
Отбор для погребения в жилищах только детей до года также указывает на бытование у населения специфического обряда. Полученные нами генетические доказательства отсутствия девочек среди восьми мла- денцев свидетельствуют об осознанном выборе мальчиков для захоронения в жилищах.
На городище Чича-1 в трех жилищах находились парные погребения младенцев. Установив, есть ли родственные связи внутри таких пар, можно выяснить, что лежит в основе обряда и его сущность. У детей, погребенных по двое в жилищах № 10 и 20, мтДНК представлена разными гаплотипами. Следовательно, младенцы не являются родственниками по материнской линии. Эти данные не исключают родства погребенных по отцовской (мужской) линии. Однако для проверки последнего предположения необходим дополнительный анализ маркеров Y-хромосомы.
Рассматриваемый обряд не был характерен для древнего автохтонного населения Западной Сибири [Там же], хотя отдельные захоронения детей на поселениях известны [Молодин и др., 2003]. Такие погребения отмечены, например, на памятнике елунинской культуры Березовая Лука на Алтае [Кирюшин, Тишкин, Грушин, 1999]. Возможно, обряд был заимствован у древних жителей Средней Азии. У них эта традиция бытовала еще с IV тыс. до н.э. [Литвинский, 1952]. В земледельческих культурах Востока захоронения детей совершались под полом, стенами и порогом жилища [Ghirshman, 1954, p. 30]. Погребения младенцев в жилищах являются характерной чертой петровской культуры Северного Казахстана (начало эпохи развитой бронзы) [Зданович, 1988, с. 133], территориально и хронологически близких им памятников нуртайского типа в Центральном Казахстане [Ткачев, 1999, с. 22–24]. Таким образом, рассматриваемый обряд тяготеет к юго-западным от Барабинской лесостепи территориям [Молодин и др., 2003, с. 315].
Данное предположение подтверждается результатами генетического анализа структуры ГВС I и II мтДНК младенцев из жилищ городища Чича-1. Гапло-типическое разнообразие мтДНК младенцев (табл. 2) в некоторой степени отражает высокую гетерогенность митохондриального генофонда всего населения Чичи-1. Выявленные структурные варианты хотя и принадлежат к кластеру западно-евразийских гап-логрупп мтДНК, но отличаются по времени и месту возникновения на континенте, а также по распространенности в современных популяциях человека.
Наиболее представительными в исследованной группе оказались линии кластера UK, объединяющего гаплогруппы U и K (образцы № 1–4). Су-пергаплогруппа U является одной из древнейших гаплогрупп западно-евразийского кластера. Ее возраст составляет свыше 50 тыс. лет [Richards et al., 1998]. Линии гаплогруппы U занимают второе место по распространенности в генофондах европеоидных популяций западной части Евразии и уступают лишь вариантам гаплогруппы H. В составе супергаплогруппы U выделяют несколько подгрупп
(U1–8), эволюционно различающихся и пространственно разобщенных в современных популяциях человека [Maca-Meyer et al., 2001].
Три из шести исследованных образцов мтДНК входят в состав супергаплогруппы U. МтДНК образца № 2 с гаплотипом 16183А→С-16189-16249 относится к гаплогруппе U1a. В современных популяциях человека гаплогруппа U1a распространена главным образом на Ближнем Востоке (друзы (Израиль) – 6,7 %, курды – 3,8, турки – 3,2 %) и Кавказе (адыгейцы – 6,0 %, северные осетины – 3,5 %) [Macaulay et al., 1999; Richards et al., 2000]. В генофондах популяций Европы линии этой группы встречаются редко, в единичных случаях. Исключение составляют популяции Центрального Средиземноморья – 3,8 % [Di Rienzo, Wilson, 1991; Francalacci et al., 1996; Torroni et al., 1998; Richards et al., 2000]. В качестве минорного компонента линии гаплогруппы U1a присутствуют в генофондах многих современных средне- и центральноазиатских популяций [Kolman, Sambuughin, Bermingham, 1996; Comas et al., 1998, 2004]. Варианты гаплогруппы U1a полностью отсутствуют в генофондах современного коренного населения Южной Сибири [Derenko et al., 2003], финно-угорских и самодийских народов Западной Сибири [Derbeneva et al., 2002; Дербенева и др., 2002; Губина, Осипова, Виллемс, 2005], а также финно-угров и тюркоязычных этносов Волго-Уральского региона [Бермишева и др., 2002].
Специфическое распространение гаплогруп-пы U1a в современных популяциях позволяет объяснить ее присутствие у населения Чичи-1 миграцией в Барабинскую лесостепь древних групп населения с юга и/или юго-запада, вероятно, с территории современного Казахстана. Это подтверждается и археологическими данными [Молодин, 2006а, б]. Действительно, варианты гаплогруппы U1a были обнаружены у представителей древнего населения Казахстана переходного от бронзы к железу периода [Lalueza-Fox et al., 2004]. Кроме того, они присутствуют в генофонде населения Синьцзяна эпохи развитой бронзы (~1 800 лет до н.э.) [Цуй Иньцю, 2003]. Кстати, последнее также может быть обусловлено связанной с миграцией в Барабу с территории современного Центрального Казахстана сначала андроновцев [Кузьмина, 1994; Molodin, Komissarov, 2004], а затем носителей бегазы-дандыба-евской культуры [Молодин, 1998].
Наличие «пришлого» компонента отмечено также присутствием в генофонде населения Чичи-1 линии гаплогруппы U3. Вариант с нуклеотидной заменой в положении 16343, относящийся к этой группе, ранее был выявлен у взрослой женщины, погребенной на территории городища Чича-1 [Молодин и др., 2006]. Картины распространения гаплогрупп U3 и U1a в современных популяциях человека в общих чертах схожи. Вероятно, присутствие гаплогруппы U3 в ге- нофонде населения городища Чича-1 также связано с привнесением ее вариантов из древних среднеазиатских или ближневосточных популяций.
Образец мтДНК № 4 со структурой гаплотипа 16356-16362 относится к гаплогруппе U4. Эта группа распределена в современных популяциях человека иначе, чем рассмотренные выше гаплогруппы U1a и U3. Ее частота возрастает с запада на восток. Несмотря на то, что гаплогруппа U4 присутствует в генофондах многих популяций Ближнего Востока [Richards et al., 2000; Metspalu et al., 2004; Di Rienzo, Wilson, 1991], ее частота здесь не превышает 3 % (сирийцы – 2,9 %) [Richards et al., 2000]. В Европе ее частота несколько выше и достигает максимума 5,4 % в популяциях Северо-Восточной Европы [Sajantila et al., 1995, 1996; Richards et al., 1998, 2000; Tolk et al., 2000]. С наибольшими частотами линии этой группы присутствуют в генофондах финно-угорских и тюркских народностей Волго-Уральского региона (максимальные частоты в этом регионе: коми зыряне – 24,2 %, чуваши – 16,4 %) [Бермишева и др., 2002], а также финно-угорских и самодийских народностей Западной Сибири и примыкающих к ней территорий Восточной Сибири: манси – 16,3 %, ханты – 18,6, нганасаны – 20,8 и кеты – 28,9 % [Derbeneva et al., 2002; Дербенева и др., 2002; Губина, Осипова, Виллемс, 2005]. На юге Западной Сибири гаплогруппа U4 отмечена у алтайцев и хакасов [Derenko et al., 2003; Дамба и др., 2003]. В среднеазиатских популяциях она встречается с небольшой частотой [Comas et al., 1998, 2004]. Таким образом, максимальными частотами линий гап-логруппы U4 характеризуются финно-угорские и самодийские народы Западной Сибири.
Гаплотип мтДНК, маркируемый мотивом 1635616362 (образец № 4), является предковым для одной из групп вариантов гаплогруппы U4, распространенных в Западной Сибири. Так, в генофондах манси и хантов распространена группа линий, характеризуемых мотивом 16113С-16356-16362, возраст которой оценивается в ~19 тыс. лет [Малярчук, 2004].
Следовательно, обнаруженный вариант гаплогруп-пы U4, по всей видимости, длительное время присутствовал на данной территории и его носителя следует отнести к автохтонной части древнего населения Ба-рабинской лесостепи. Примечательно, что гаплотип гаплогруппы U4 выявлен у младенца, погребенного в жилище на территории «цитадели» – секторе городища, где проживало предположительно аборигенное для Барабы население.
Образец мтДНК № 3 с гаплотипом 16189-1626016270 относится к гаплогруппе U5b. Линии этой группы распространены по всей территории Европы, до стигают частот ок. 5 % в популяциях Северо-Восточной Европы (без учета популяции саамов) [Sajantila, 1995, 1996; Richards et al., 2000]. Сопоста- вимые частоты линий гаплогруппы U5b характерны для популяций Волго-Уральского региона [Бермишева и др., 2002]. В популяциях Ближнего Востока, Кавказа и Средней Азии присутствуют лишь ее единичные линии. На территории Западной Сибири гаплогруп-па U5b выявлена в митохондриальном генофонде алтайцев с частотой 2,7 % [Derenko et al., 2003]. В генофондах финно-угорских и самодийских народностей севера Западной Сибири варианты гаплогруппы U5b не обнаружены.
Гаплогруппа K, к которой отно сится образец мтДНК № 1 с гаплотипом 16093-16224-16311, широко распространена в популяциях западной половины Евразийского материка. Линии гаплогруппы K представлены в генофондах большинства популяций Европы, Ближнего Востока и Кавказа с частотой от 3 до 10 % (в среднем ок. 6 %) [Macaulay et al., 1999; Richards et al., 2000]. С меньшей частотой они встречаются в популяциях Средней Азии, а также Волго-Уральского региона [Comas et al., 1998, 2004; Бермишева и др., 2002]. В современных популяциях Западной Сибири немногочисленные линии этой гаплогруппы присутствуют только в генофондах финно-угорских народов.
МтДНК образца № 6 содержит нуклеотидные замены в положениях 16069-16126, что позволяет однозначно определить ее принадлежность к западно-евразийской гаплогруппе J [Richards et al., 1998]. Гаплогруппа J возникла порядка 45 тыс. л.н. на Ближнем Востоке; здесь выявлено наибольшее разнообразие линий этой гаплогруппы. Современные ближневосточные популяции характеризуются самыми высокими частотами этой гаплогруппы в митохондриальных генофондах – в среднем 12 % [Richards et al., 2000; Metspalu et al., 2004]. В некоторых популяциях этого региона ее частота превышает 20 %: у бедуинов Аравийского полуострова – 25 % [Di Rienzo, Wilson, 1991], у арабов Саудовской Аравии – 22,5 % [Abu-Amero et al., 2007]. В европейских популяциях частота гаплогруппы J несколько ниже – порядка 10 % [Richards et al., 2000]. С близкой частотой она встречается в популяциях Кавказа – в среднем менее 8 % (максимум у северных осетин – 9,6 %) [Macaulay et al., 1999; Richards et al., 2000; Comas et al., 2000; Bermisheva et al., 2004]. В качестве минорного компонента линии гаплогруппы J присутствуют в генофондах популяций Средней и Центральной Азии, а также юга Западной Сибири [Derenko et al., 2003]. У финно-угров Западной Сибири частота гаплогруп-пы J существенно выше и сопоставима с европейским и ближневосточным уровнями (манси – 12,2 %, ханты – 11,9 %). Структура ГВС I образца мтДНК № 6 совпадает с вариантом-основателем гаплогруппы J. Известно, что гаплотипы с идентичной структурой ГВС I могут различаться заменами нуклеотидов на других участках мтДНК. Определив структуры этих участков ДНК, в дальнейшем можно уточнить дополнительную филогенетическую информативность обнаруженного варианта мтДНК.
В ГВС I образца мтДНК № 7 выявлена всего одна нуклеотидная замена в положении 16366. По-види-мому, этот структурный вариант является непосредственным производным кембриджской референсной последовательности и относится к западно-евразийской гаплогруппе H. Гаплогруппа Н с наибольшей частотой встречается у населения Западной и Северной Европы (40–50 %), с частотой 20–40 % – Южной, Юго-Западной, Восточной Европы, с частотой менее 20 % – Ближнего Востока и Индии [Torroni et al., 1996]. С довольно высокой частотой она присутствует в митохондриальных генофондах финно-угорских народов Западной Сибири (манси – 14,3 %, ханты – 19, коми – 26,9 %) [Губина, Осипова, Виллемс, 2005; Derbeneva et al., 2002], что сближает их с популяциями Европы и Ближнего Востока. Высокая частота вариантов гаплогруппы Н отмечена у шорцев (21,4 %) [Деренко и др., 2001]. С меньшими частотами линии группы Н присутствуют в генофондах самодийцев (кеты – 10,5 %, нганасаны – 8,4 %) и некоторых народностей юга Западной Сибири. В составе гапло-группы H выявлено большое число подгрупп (свыше 15) [Loogvali et al., 2004]. Высокая информативность некоторых субгаплогрупп Н проявляется при фи-логеографических исследованиях популяций человека [Achilli et al., 2004]. Поэтому оперирование в анализе только сведениями, касающимися структуры последовательности нуклеотидов ГВС I мтДНК в случае гап-логруппы H, малоинформативно. Для детального фи-логеографического анализа требуется установление конкретной подгруппы [Roosralu et al., 2007].
Итак, несомненный интерес представляет тот факт, что варианты мтДНК относительно небольшой выборки младенцев, погребенных в жилищах городища Чича-1, принадлежат к тем западно-евразийским гаплогруппам, значительная часть которых отсутствует либо представлена с незначительными частотами в генофондах современных групп населения региона. Наряду с линиями мтДНК, ныне распространенными среди жителей территорий, которые расположены южнее и юго-западнее Барабы (Казахстан, Средняя Азия, Ближний Восток, Кавказ), в анализируемой выборке выявлены также носители предковых вариантов гап-логрупп, распространенных среди современного коренного населения Западной Сибири.
Провести полноценное сравнение особенностей генофонда мтДНК групп, населявших центральную и периферическую части территории городища, пока не представляется возможным ввиду небольшой численности исследованной выборки. Для этого необходимо проанализировать дополнительные образцы ДНК из палеоантропологических материалов того времени. Раскопки некрополя, отно сящего ся к городищу [Молодин и др., 2004], во сточная периферия которого сформирована мигрантами [Молодин, 2006а], позволяют надеяться, что с привлечением останков из некрополя можно будет полнее охарактеризовать генетические различия между автохтонным и пришлым населением. Несомненно, требуется изучение структуры не только последовательностей нуклеотидов контрольного района, но и других фрагментов мтДНК. На данном этапе работы, используя генетические данные, мы можем констатировать присутствие на городище смешанного – автохтонного и пришлого – населения, что согласуется с выводами, полученными при анализе археологических источников.
