Палеогеографические особенности развития растительности и климата Северо-Западного Алтая в плейстоцене

Автор: Болиховская Н.С., Шуньков М.В.
Журнал: Археология, этнография и антропология Евразии @journal-aeae-ru
Рубрика: Палеоэкология. Каменный век
Статья в выпуске: 2 (58), 2014 года.
Бесплатный доступ
На основе данных, полученных при детальном палинологическом изучении плейстоценовых отложений многослойных палеолитических стоянок Северо-Западного Алтая, реконструированы последовательности климатических, флористических и фитоценотических изменений, происходивших в межледниковые и холодные этапы плейстоцена в низкогорной части Алтайских гор, в периоды обитания здесь палеолитического человека. Установлены палеогеографические особенности оптимумов изученных неоплейстоценовых межледниковых эпох и пессимумов периодов похолоданий ледникового ранга на этой территории.
Неоплейстоцен, палинология, палеогеография, природная среда, растительные формации, структура природных комплексов
Короткий адрес: https://sciup.org/14523043
IDR: 14523043 | УДК: 551.891(235.222)+903.3
Текст научной статьи Палеогеографические особенности развития растительности и климата Северо-Западного Алтая в плейстоцене
Неполнота геологической летописи плейстоцена на большей части Северной Евразии обусловливает необходимость выявления особенностей пространственно-временных закономерностей развития флоры, растительности и климата для решения вопросов детального расчленения новейших отложений, периодизации и корреляции климатообусловленных палеогеографических событий, а также определения геологического возраста палеолитических объектов и реконструкции условий обитания первобытного человека. Материалы по истории растительного покрова входят в число важнейших палеогеографических свидетельств, на основе которых К.К. Марков сделал вывод о том, что главными закономерностями изменения природной среды являются направленность, ритмич- ность и метахронность (местная индивидуальность) [1960]. Накопление и синтез новых аналитических данных в ходе последующих палеогеографических исследований позволили вне сти существенные коррективы в представления об особенностях трех указанных пространственно-временных закономерностей развития флоры, растительности и климата плейстоцена [Болиховская, 2005, 2007].
На основе детальных реконструкций флористических, фитоценотических и климатических сукцессий, созданных для ряда страторайонов Восточной Европы, которые имеют различную историю палеогеографического развития, уточнены структура климатической ритмичности – количество межледниковых и ледниковых ритмов в пределах неоплейстоцена – и особенности климаторитмики внутри теплых и холодных эпох. Установлено, что изменения природной среды на протяжении хрона Брюнес были обусловлены сменами 15 глобальных климатических ритмов – 8 межледниковий и 7 похолоданий ледникового
ранга. В пределах ледниковых и межледниковых этапов выделены более дробные климатостратиграфические единицы: в межледниковых климатических ритмах прослежены эндотермальные похолодания, термоксеротические и термогигротические стадии, а в холодных ритмах ледникового ранга – стадиалы, межстадиалы, межфазиалы, криогигротические и криоксеротические стадии.
Детальные палинологические записи значительно расширили наши знания о специфике пространственной дифференциации растительного покрова межледниковых и ледниковых эпох. Пространственная дифференциация перигляциальной растительности, т.е. растительности ледниковых периодов неоплейстоцена, была более значительной, чем пространственная дифференциация растительного покрова межледниковых эпох. Разнообразие зональных и подзональных типов растительности холодных периодов было бóльшим, чем разнообразие зональных и подзональных типов растительности межледниковых эпох [Бо-лиховская, 2005].
Полученные материалы существенно изменили также представление о ходе направленного развития флоры и растительности в плейстоцене. Они подкрепили лишь повсеместно прослеживаемую закономерность направленного обеднения всей совокупности межледниковых флор одного звена плейстоцена, по сравнению с интегральными межледниковыми флорами последующего звена, установленную по результатам исследований позднекайнозойских флор высших растений во всех изученных районах Северной Евразии. Однако они не подтвердили господствующее мнение о том, что каждая последующая межледниковая эпоха должна характеризоваться флорой, более бедной экзотическими элементами, чем флора предшествующего межледниковья. Согласно полученным результатам, нараставший в позднем кайнозое процесс обеднения межледниковых флор экзотическими элементами в отдельные межледниковые эпохи раннего и среднего неоплейстоцена нарушался появлением флор с более разнообразным составом таксонов и более богатым набором неогеновых реликтов, чем флора предшествующего межледниковья [Болихов-ская, 2007].
Кроме того, установлена еще одна закономерность в истории растительности и климата неоплейстоцена. Путем сравнительного анализа почти непрерывной последовательности климатофитоценотических и флористических сукцессий определены два длительных цикла изменения флоры, растительности и климата Восточно-Европейской равнины в неоплейстоцене, обусловливавшие природные особенности всех межледниковых и холодных этапов. Каждый из этих циклов охватывал четыре межледниковые и четыре ледниковые эпохи [Болиховская, 2005].
Данные по абсолютной хронологии теплых и холодных эпох позволили установить возрастной диапазон каждого такого цикла. Для определения возраста и продолжительности реконструированных теплых и холодных этапов неоплейстоцена проведена корреляция континентальных отложений и палеоклиматиче-ских событий внеледниковой и ледниково-перигляциальной зон Восточно-Европейской равнины с теплыми климатическими ритмами, реконструированными на основе ЭПР-анализа раковин морских моллюсков из трансгрессивных отложений палеошельфовой зоны Северной Евразии, и с изотопно-кислородной шкалой [Болиховская, Молодьков, 2002; Bolikhovskaya, Molodkov, 2006; Molodkov, Bolikhovskaya, 2006]. Согласно этим хроностратиграфическим данным, продолжительность установленных циклов в развитии природной среды составляла примерно 450 тыс. лет. Каждый из межледниковых или ледниковых этапов более молодого 450-тысячелетнего цикла имел свой палеогеографический аналог в предшествовавшем цикле. Открытие этой закономерности вывело на новый уровень перспективы палино-климатострати-графических исследований в решении вопросов стратиграфии и палеогеографии плейстоцена не только равнинных, но и горных областей.
Одной из наиболее изученных в этом отношении горных областей Северной Евразии является территория Российского Алтая, входящая в состав Евроазиатской степной зоны. При этом северная часть Алтая зонально находится в подзоне северной лесостепи, связанной через леса Кузнецкого Алатау с южной тайгой, а южная часть примыкает к зоне северных пустынь Центральной Азии. Кроме того, флорофито-ценотическая специализация этого района обусловлена также тем, что Алтай входит в обширную зону контакта Бореальной (отно сительно гумидной) и Центрально-азиатской (аридной) областей Голаркти-ки [Толмачев, 1974]. По территории Алтая проходит граница между двумя секторами Северной Азии – бореальным Западносибирско-Среднеазиатско-Гималайским и экстремоконтинентальным Централь-носибирско-Центральноазиатско-Индокитайским [Камелин, 2005]. Эти закономерности широтной зональности и секторальности накладывают отпечаток на особенности современной вертикальной поясности Северо-Западного Алтая, где сейчас распространены сообщества четырех высотных поясов: высокогорного (с альпийско-луговыми и горно-тундровыми ассоциациями); горно-таежного; переходного горно-лесостепного, объединяющего горно-степные и горно-таежные ассоциации; горно-степного [Огуреева, 1980].
Необходимо отметить, что в настоящее время широколиственные и хвойно-широколиственные лесные формации в составе растительности Алтая отсутствуют. Более того, здесь не представлены почти все немо- ральные древесно-кустарниковые растения, автохтонная пыльца которых обнаружена в плейстоценовых отложениях этой горной страны. Из всех широколиственных пород, входящих в состав ископаемой плейстоценовой дендрофлоры района исследований, только липа сибирская Tilia sibirica встречается на северо-востоке Алтая. Вместе с тем на юго-западе и северо-востоке Алтая есть районы с довольно высоким среднегодовым количеством осадков (от 1 000 до 2 000 мм), сохранившие в современном растительном покрове травянистые реликты – спутники широколиственных лесов и даже формации – дериваты хвойно-широколиственных лесов плиоцена [Куминова, 1957]. На их территории находились главные алтайские рефугиумы неморальных лесных формаций, не прекращавшие своего существования в самые неблагоприятные в климатическом отношении периоды плейстоцена.
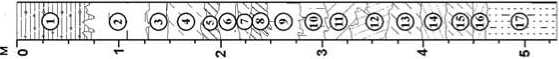
Имеющаяся сегодня палеоботаническая информация для отдельных районов Алтая различна по объему. Наиболее представительные климатофло-рофитоценотические записи, позволяющие выявить специфические черты изменения плейстоценовой флоры и растительности, получены для северо-западной части Алтайских гор [Природная среда…, 2003]. Благодаря долгосрочным междисциплинарным исследованиям многослойных палеолитических объектов, расположенных в долине р. Ануй, – Карама, Денисова пещера, Усть-Каракол, Ануй-2 и др. (рис. 1), постоянно пополняются геологические, палеопочвенные, палеоботанические, палеофаунистические, геохронологические и другие аналитические данные, необходимые для обоснования возраста и климатостратиграфического расчленения плейстоценовых отложений, воссоздания истории развития природной среды и условий обитания первобытного человека.
Палеоботанические исследования новейших отложений на этой территории проводятся с начала 1990-х гг. В 1992–1997 гг. Е.М. Малаева выполнила детальное спорово-пыльцевое исследование неоплей-стоценовых толщ палеолитических стоянок Денисова пещера, Усть-Каракол, Ануй-2, разрезов Нижний Каракол и Черный Ануй, расположенных ныне в горно-таежном поясе. Был проведен скрупулезный историкофлористический анализ изученных плейстоценовых палинофлор и подробно реконструированы изменения растительности и климата в период от тобольского межледниковья до голоцена [Малаева, 1995, 1998; Деревянко, Малаева, Шуньков, 1998, 2000]. Кроме того, Е.М. Малаева дала оценку двум теплым этапам господства лесных и лесостепных фитоценозов в раннем неоплейстоцене [Деревянко и др., 1992].
С 1998 г. палинологические исследования плейстоценовых отложений Северо-Западного Алтая проводятся Н.С. Болиховской. К настоящему времени на основании результатов спорово-пыльцевого анализа разрезов стоянок Карама, Каминная пещера и др. реконструированы изменения растительности и климата, происходившие на протяжении двух межлед-
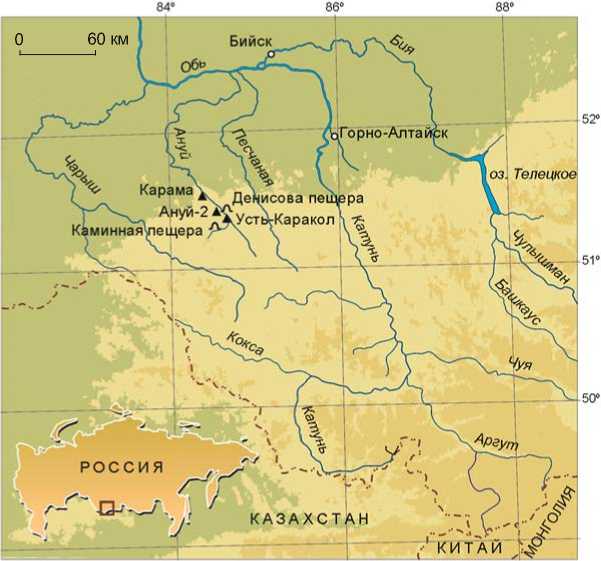
Рис. 1. Опорные палеолитические комплексы в бассейне р. Ануй, Северо-Западный Алтай.
никовых и двух холодных эпох раннего неоплейстоцена, которые коррелируют с морскими изотопными стадиями 16–19 теплого каргинского этапа, пессимума сартанского времени и позднеледниковых сартанских стадиалов и межстадиалов [Болиховская, Маркин, 2002; Бо-лиховская, Шуньков, 2005; Деревянко и др., 2000; Bolikhovskaya, Derevyanko, Shunkov , 2006].
Нижний неоплейстоцен
Материалы по климатостратиграфии и флорофитоценотическим реконструкциям, полученные для нижненеоплей-стоценовых этапов развития природной среды Северо-Западного Алтая, имеют большое значение для установления закономерностей изменения растительности и климата не только Алтая, но и всей южной территории Северной Азии.
Осадки нижнего неоплейстоцена в долине Ануя впервые изучены в разрезах Черный Ануй и Нижний Каракол,
расположенных в пределах современного горно-лесного пояса на абсолютных высотах 740–750 м [Деревянко и др., 1992]. Спорово-пыльцевые спектры этих разрезов характеризуют растительность двух ранне-неоплейстоценовых теплых эпох, в составе дендрофлоры которых наряду с эдификаторами современных лесов произрастали экзоты: Betula sect. Costatae , Juglans cf. mandshurica , Carpinus betulus , Quercus , Ulmus cf. laevis , Tilia sibirica , Acer , Alnus , Corylus avellana . Во время первого межледниковья в окрестностях разреза Черный Ануй были развиты лесостепи с господством березовых лесов и участием сосны обыкновенной, кедра сибирского и широколиственных пород. Последующее похолодание с увеличением влагообеспеченности привело к доминированию лесных сообществ из Pinus sylvestris , P. sibirica , Abies и Picea . Вторая межледниковая эпоха отражена в пали-носпектрах разреза Нижний Каракол. На протяжении этой эпохи сначала господствовали леса, затем лесостепи, а позднее вновь заметно возросло участие лесных сообществ. В оптимальные фазы, как и во время первого потепления, преобладали березовые леса с участием широколиственных и хвойных деревьев.
Более полные и детальные палинологические данные, характеризующие не только теплые, но и холодные эпохи нижнего неоплейстоцена, получены при изучении разреза раннепалеолитической стоянки Ка-рама, расположенной на абсолютной высоте ок. 630 м. Судя по геоботаническим описаниям растительного покрова Алтая [Огуреева, 1980, с. 5] и структуре фитоценозов в окрестностях стоянки, она находится в настоящее время в переходной зоне от горно-лесного к горно-лесостепному поясу.
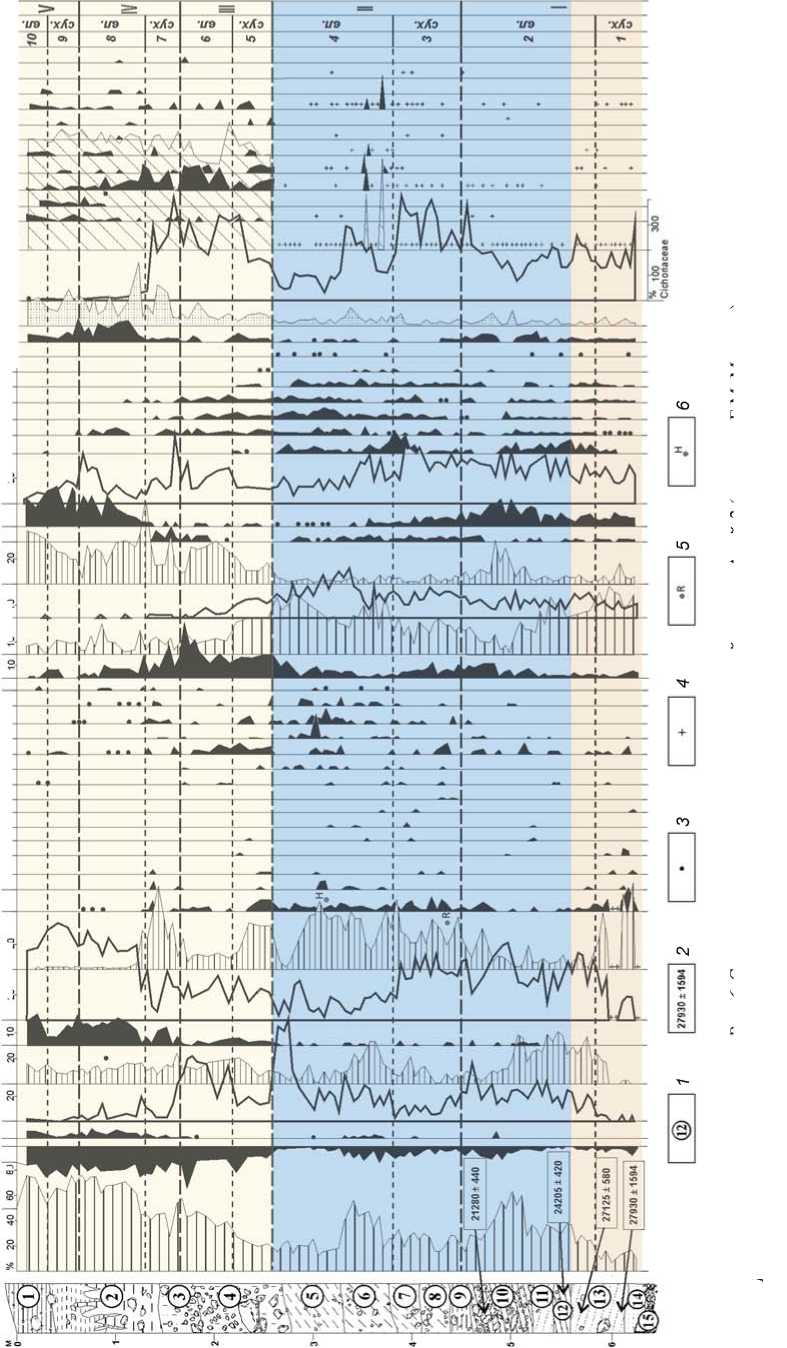
Раскоп Карамы вскрыл толщу осадков общей мощностью ок. 12 м, которая, согласно характеру залегания, структуре осадка и динамике палиноспектров, состоит из трех толщ и имеет значительные седиментационные перерывы [Стоянка…, 2005]. Детальный спорово-пыльцевой анализ вскрытых отложений позволил уточнить их геологический возраст, реконструировать ландшафтно-климатические условия обитания раннепалеолитического человека, подробно охарактеризовать изменения флоры и растительности, происходившие на протяжении древнейших межледниковых и ледниковых эпох неоплейстоцена [Болиховская, Шунь-ков, 2005; Bolikhovskaya, Derevyanko, Shunkov , 2006].
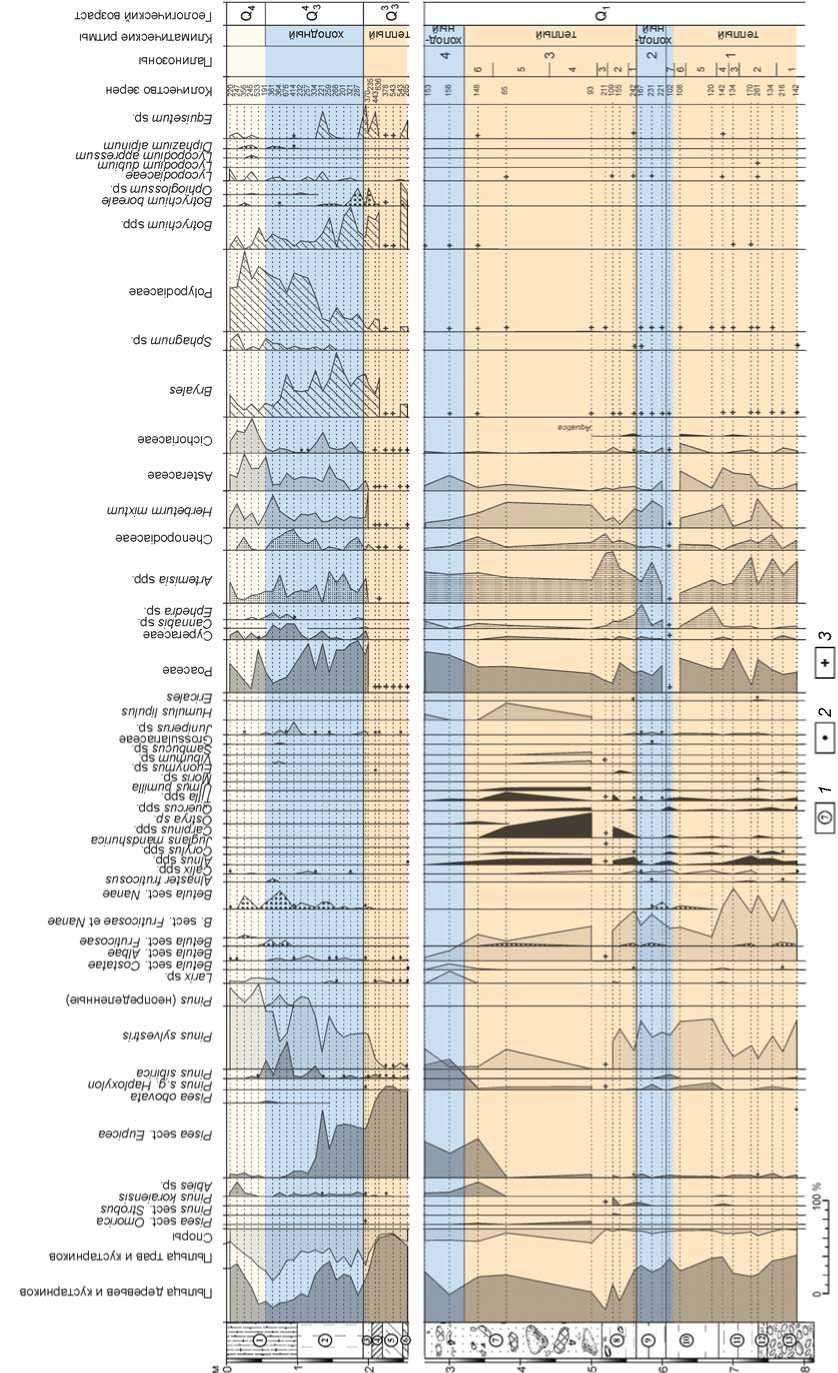
В целом в составе автохтонной палинофлоры трех проанализированных толщ Карамы отмечено более 130 таксонов разного ранга. На приведенной ниже спорово-пыльцевой диаграмме (рис. 2) большинство определенных до вида таксонов объединены и показаны в составе родов и семейств. Определена пыльца 45 родов и видов деревьев и кустарников, а также 56 видов, родов и семейств травяно-кустарничковых растений. Группу высших споровых растений
(мхов, папоротников, плаунов, хвоща и др.) представляют 30 таксонов.
Палинофлора нижней (слои 13–9) и средней (слои 8 и 7) толщи разреза Карама существенно богаче, чем флоры изученных ранее одновозрастных отложений разрезов Черный Ануй и Нижний Каракол, содержавших только 25 родов и видов древесно-кустарниковых таксонов. Вместе с тем по родовому и видовому составу дендрофлоры, а также по структуре палеофитоценозов периоды накопления этих отложений относительно близки к древним межледниковым эпохам, реконструированным для разреза Карамы.
Палиноспектры слоев 7, 8 и 10–13 Карамы включают значительное количество экзотических для современной дендрофлоры элементов, принадлежащих бореальным: Picea sect. Omorica, Pinus sect. Strobus, Pinus cf. koraiensis, Betula sect. Costatae и неморальным европейским, дальневосточным и другим таксонам: Juglans mandshurica , Carpinus betulus , C. cordata , C. orientalis , Ostrya sp., Quercus robur , Tilia cordata , T. amurensis, T. mandshurica , Ulmus pumila , Corylus avellana , Alnus glutinosa , A. incana , Morus sp. и др. Многие из них впервые отмечены в палиноспектрах средне- и ранненеоплейстоценовых отложений долины Ануя.
Присутствие в палинофлоре слоев 7, 8 и 10–13 Pinus sect. Strobus, Carpinus cordata, C. orientalis, Ostrya sp. , Quercus robur, Tilia cordata, T. amurensis, T. mandshurica, Alnus glutinosa , A. incana , Corylus avellana , Juglans mandshurica , Carpinus betulus , Ulmus pumila, Morus sp. и др., а также эколого-ценотические особенности всех обнаруженных экзотических таксонов, результаты анализа географических групп родов дендрофлоры и другие данные свидетельствуют о ранненеоплейстоценовом возрасте этого разреза. Например, хмелеграб Ostrya sp. и шелковица Morus sp . , принадлежащие к группе американо-средиземноморско-азиатских родов, являются показателями ранне-неоплейстоценового возраста вмещающих отложений Южного Прибайкалья и Верхнего Приамурья [Махова, 1978; Гричук, 1982].
Межледниковые палинофлоры Карамы существенно отличаются от эоплейстоценовых флор Восточного Алтая, юга Западной Сибири и других горных и равнинных областей Южной Сибири и Дальнего Востока [Гитерман и др., 1968; Волкова, 1977; Голубева, Караулова, 1983; Белова, 1985; Архипов, Волкова, 1994]. В них отсутствуют пыльцевые зерна субтропических широколиственных пород: Pterocarya , Carya , Zelkova, Celtis, Ilex , тсуги Tsuga и других экзотических таксонов семейства сосновых, характерных для термофильных эоплейстоценовых флор указанных районов. Это обстоятельство не дает оснований предполагать для отложений Карамы эоплейстоценовый возраст [Боли-ховская, Шуньков, 2005]. Вместе с тем некоторые ис-
:5s2Zrsr dds eisiiua^jv ds ejpaqdg ds siqeuueo aeaoejadXo eaoidng joas easy
-t/ouox
HAuuai
H«l " l»H ~ I - еэцепйу.
ds iunssoj6oiqdo a/easoq ujniqoAjiog
Vouox
HRuuai
* H N H j з s g ? 5 eg 5
loedsoe nnxaahnjouoaj iami nd аихэакигемиих aedoeipodApd ds шnu6eчds saieAjg aeaoeuoqoiQ aeaoejaisv шп)х>ш шJn|aqJЭн aeaoeipodouaqo s^sa/qAs snuy eouiqis snuy uo/Axo/deH 6 s snuy е^елоро easy aoxuHdeioA* и aaqeadaV ehquiqu
HRHtfOUOX
HRUUdl aeaoeod sajeofjy Snindl/ 5П{ПШПН ds sruadiunr aeaoeueinssojg ds sninqwes ds tunwnqiA ds snujAuonq ds S1JO№ eiifiundsmurn dds snojano ds gAjiso dds snuidjeo еоипцзри^ш sue^n?
odssnuw dds xqeo snsooiinjj jaiseujy aeueN toas ein^ag aeueN ia aesooqny pas g
(atAHHauaVaduodH) snuy ds saiqv sisuaiex»! snuy snqojig' toes snuy eoixxuQ joas easy lAdOUQ eoxuHdeioXx и eedi еЬяшчи aesooqnj-j aeqiv ae;eisoo pas einiag pas e/niag pas einiag ds xuei
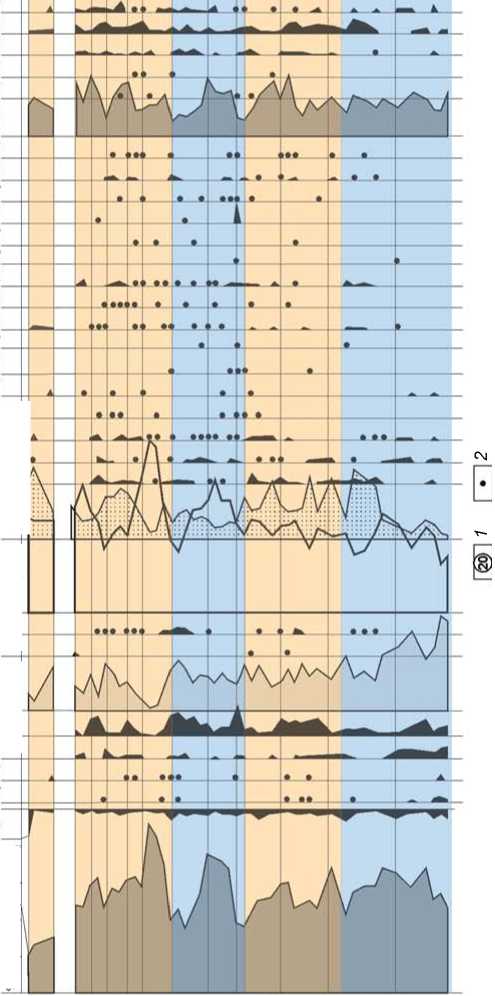
Рис. 2. Спорово-пыльцевая диаграмма отложений стоянки Карама (аналитик Н.С. Болиховская).
1 - литологический слой; 2 - содержание пыльцы менее 1 %; 3 - наличие спор в малочисленной группе (без подсчета процентов).
следователи считают, что для нижней толщи Карамы нельзя исключать эоплейстоценовый возраст [Зыкин и др., 2005]. Это предположение основано на палео-магнитном тестировании разреза, показавшем для всей толщи отложений прямую намагниченность, а также присутствие в строении нижней части разреза педокомплекса из двух слитоземных почв, которые до этого были известны только в плиоценовых отложениях Западной и Средней Сибири, что, по мнению В.С. Зыкина и его коллег, допускает корреляцию низов караминского разреза с барнаульскими слоями эоплейстоцена и, соответственно, с субхроном Олду-вей. Однако с этим предположением трудно согласиться, поскольку ранненеоплейстоценовые межледниковые эпохи, реконструированные по палинологическим данным Карамы, по флористическим, фитоценотиче-ским и палеоклиматическим характеристикам заметно отличаются от периода формирования барнаульских слоев на юге Западной Сибири [История…, 1970]. Если отложения с барнаульской флорой накапливались в лесостепях и степях при климате, близком к современному [Волкова, Кулькова, 1999], то межледниковые флоры Карамы существовали в значительно более теплых и менее континентальных климатических условиях, чем современные.
Спорово-пыльцевые спектры Карамы снизу вверх по разрезу отражают значительную динамику состава и процентного содержания таксонов, свидетельствующую о неоднократных сменах зональных типов растительности в долине Ануя и перегруппировках растительных формаций в окрестностях стоянки, обусловленных глобальными климатическими колебаниями четырех реконструированных крупных этапов хрона Брюнес.
Время первого зафиксированного в разрезе межледниковья соответствует периоду накопления толщи слоев 13–10. Эта эпоха сопоставлена с морской изотопной стадией (МИС) 19, или гремячьев-ским межледниковым этапом Восточно-Европейской равнины, который по корреляционным подсчетам датируется интервалом 790–760 тыс. л.н. [Болиховская, 2005; Molodkov, Bolikhovskaya, 2009]. В это время в более теплом, чем современный, климате доминировали лесостепные и лесные ландшафты. Реконструированы семь фаз, отражающих трансформации составлявших их сообществ – участков степей, широколиственных лесов из Tilia cordata и T. sibirica, Quercus robur, Q. mongolica Fisch. ex Ledeb., Carpinus cordata, Juglans mandshurica, Ulmus, Morus, березовых и хвойных лесов. Фаза 1 (палинозона 1.1) – господство сосново-березовых и березово-сосновых лесов из Betula pendula, B. pubescens, Pinus sylvestris с примесью ели, Betula sect. Costatae, дуба и липы Tilia cordata, T. sibirica, с Corylus avellana в подлеске. Пыльца термофильных элементов дендрофлоры в сумме составляет 3,5–7,9 %. Фаза 2 (палинозона 1.2) – доминирование лесостепи. Доля пыльцы неморальных видов дендрофлоры возросла до 9,2 %. В условиях потепления климата сократилась общая площадь лесов, но в их составе повысилась роль широколиственных пород: Carpinus cordata, Quercus robur, Tilia cordata, T. sibirica, Ulmus pumila, Morus и др. В степных биотопах сначала преобладали злаки и разнотравье, а затем марево-полынные и астрово-полынные сообщества. Фаза 3 (палинозона 1.3) отвечает эндо-термальному (внутримежледниковому) похолоданию, в течение которого по-прежнему господствовали лесостепи. На снижение теплообеспеченности указывает резкое падение (до 1,4 %) роли пыльцы широколиственных пород, представленной только Tilia sibirica. В степных группировках стали превалировать Poaceae, Asteraceae, Artemisia. Фаза 4 (палинозо-на 1.4) отражает увеличение гумидности климата и господство лесных ландшафтов – содержание пыльцы древесных пород увеличилось до 70 %. Доминировали сосново-березовые леса. Благодаря относительному потеплению и росту влагообеспеченности в составе лесов появились Abies, Pinus cf. koraiensis, Betula sect. Costatae, Juglans mandshurica, Quercus. Фаза 5 (палино-зона 1.5) – превалирование кедрово-сосновых и березово-дубово-липово-грабовых лесов. Фаза 6 (палинозо-на 1.6) характеризуется дальнейшей трансформацией лесной растительности при заметном снижении тепло- и влагообеспеченности во время второго эндотермала. Сократилась роль широколиственных пород, исчезли граб и дуб. Преобладали березово-сосновые леса с участием ели, Pinus cf. koraiensis, Tilia cordata, T. sibirica. В подлеске появилась кустарниковая береза. Фаза 7 (палинозона 1.7) соответствует термогиг-ротическому максимуму межледниковья. Содержание пыльцы термофильных видов дендрофлоры в сумме возросло до 14 %. Господствовали широколиственные леса из Quercus sp., Carpinus cordata, Tilia cordata, T. sibirica, Ulmus sp. с участием Alnus glutinosa. Им сопутствовали хвойно-березовые древостои из ели, Pinus sylvestris, Pinus cf. koraiensis и березы.
Формирование отложений слоя 9 (палинозона 2) происходило в холодную эпоху, сопоставленную с МИС 18, или девицким ледниковым этапом ВосточноЕвропейской равнины (760–710 тыс. л.н.). В это время были развиты перигляциальные ландшафты. В самые холодные интервалы здесь господствовали березовососновые леса, кустарниковые сообщества из Betula sect. Fruticosae, B. fruticosa, B. sect. Nanae, Alnaster fruticosus, Juniperus и др., луговые и лугово-болотные ценозы. Находки в виде единичной пыльцы липы сибирской и ольхи в палиноспектрах из подошвы и кровли слоя 9 показывают, что в подфазы, переходные от предшествующего термохрона к похолоданию и от него к следующему межледниковью, эти породы вхо- дили в состав долинных древостоев в наиболее благоприятных для них местах обитания.
Делювиально-пролювиальные осадки слоя 8 и большей части слоя 7 накапливались в следующее межледниковье (палинозоны 3.1–3.6), сопоставленное с МИС 17, или семилукским межледниковым этапом (710–660 тыс. л.н.). В климатическом отношении оно было более теплым и сухим, чем предшествующее межледниковье. Для этого термохрона реконструированы шесть фаз в развитии господствовавших степей и лесостепей. В первую фазу (палинозона 3.1) преобладали лесостепи, сочетавшие участки разнотравно-злаковых степей, широколиственных лесов из Carpinus betulus, C. cordata, Quercus sp. , Tilia mandshurica и др., сосново-березовых лесов и ольшаников из Alnus glu-tinosa и A. incana . Во вторую фазу (палинозона 3.2) доминировали степи, в составе лесных участков значительно возро сла роль широколиственных пород. Сначала преобладали грабовые из Carpinus cordata и C. orientalis и сосново-березовые сообщества. Затем повысилась влагообеспеченность и возросло значение Carpinus cordata , Tilia mandshurica , T. sibirica и темнохвойных пород. Появились Pinus sect. Strobus , P. cf. koraiensis . В третью фазу (палинозона 3.3) господствовали степи из разнотравно-злаковых и полынных сообществ, а также долинные грабовые леса из Carpinus cordata и C. orientalis с примесью Juglans mandshurica , Tilia mandshurica , Alnus glutinosa .
Со второй половиной этого межледниковья связано самое значительное расселение термофильных элементов дендрофлоры (доля их пыльцы в спектрах составляет 27–33 %). В четвертую фазу (палинозона 3.4) доминировали лесостепи, в которых основные площади занимали разнотравно-злаковые степи и парковые грабовые леса из Carpinus betulus, C. cordata и C. orientalis с примесью дуба, вяза и ольхи. Ограниченное распространение имели березовые колки. Для пятой фазы (палинозона 3.5) были характерны лесостепные ландшафты с преобладанием в составе господствовавших широколиственных лесов липово-грабовых сообществ из Carpinus cordata, Tilia cordata и T. mandshurica . В шестую фазу (палинозона 3.6) в лесостепях произошли значительные изменения в составе лесной растительности. В число доминантов вошли темнохвойные породы, в т.ч. Picea sect. Omorica . Исчезли граб и некоторые виды липы. Основные площади занимали широколиственно-пихтово-еловые леса с участием Tilia mandshurica , Ostrya sp. с примесью ольхи и берез Betula sect. Costatae, B. pendula, B. pubescens .
Отложения верхней части слоя 7 накапливались в следующую холодную эпоху (палинозона 4), сопоставленную с МИС 16, или донским оледенением (660–610 тыс. л.н.). В это время в условиях значительного похолодания климата доминировали перигляциальные степи с небольшими участками хвойного редколесья. Из лесных биотопов исчезли широколиственные породы. Преобладали лиственничные, сосновые и кедрово-еловые редкостойные леса из Larix sibirica, Pinus sylvestris, Pinus cf. koraiensis и ели. В травяно-кустарничковом покрове превалировали злаковые, разнотравно-злаковые, полынные и луговые гроздовниковые сообщества.
Средний неоплейстоцен
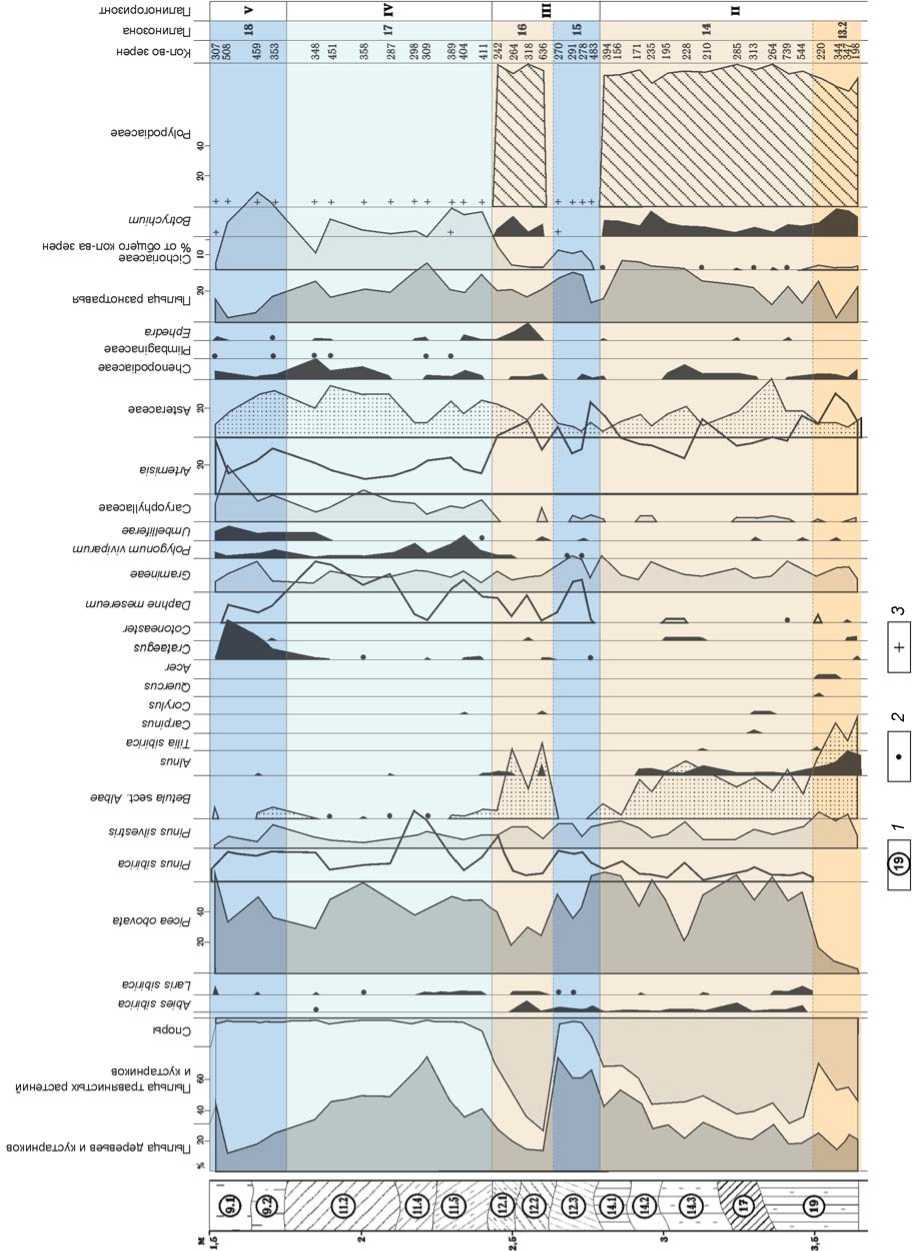
Климатофитоценотические условия обитания человека в долине Ануя в теплые и холодные эпохи среднего неоплейстоцена подробно воссозданы по палинологическим данным из отложений Денисовой пещеры (рис. 3) [Деревянко, Малаева, Шуньков, 2000]. В тобольское межледниковье (МИС 9, чекалинское межледниковье Русской равнины, 340–280 тыс. л.н.) в условиях теплого и умеренно влажного климата господствовали лесные сообщества – березовые и сосново-березовые леса с примесью ореха маньчжурского, граба обыкновенного, липы сибирской, дуба, клена, вяза гладкого, лещины, а также долинные ольховые леса с участием ели. К южным склонам долины были приурочены горно-степные травяно-кустарничковые группировки. Верхний ярус северных склонов занимали березовые и сосново-березовые леса с темнохвойными породами и лиственницей.
Во время похолодания самаровского ледникового этапа (МИС 8, калужское похолодание Русской равнины, 280–240 тыс. л.н.) существенно сократились площади лесных массивов, увеличилась доля темнохвойных пород, возросла роль степных, нивальных и бореальных видов. В заключительную фазу преобладали ксерофитные и луговые степные группировки, а небольшие лесные участки были представлены сосново-березовыми ассоциациями с участием пихты, ели, кедра и лиственницы, а также единичными вкраплениями граба, клена и лещины.
Для растительности ширтинского межледниковья (МИС 7, черепетьское межледниковье Русской равнины, 240–205 тыс. л.н.) было характерно преобладание массивов сосново-березовых лесов с примесью ели, ольхи и широколиственных пород. При этом роль по следних существенно возросла на заключительном этапе в условиях теплого и умеренно влажного климата.
В растительном покрове финальной фазы тазов-ского похолодания (МИС 6, днепровское/московское оледенение Русской равнины, 205–145 тыс. л.н.) доминировали степные и луговые сообщества из полыни, злаков и астровых. Небольшие лесные массивы включали березово-сосновые ассоциации с примесью ольхи, ели, кедра и единичных широколиственных пород – дуба, липы и вяза.
iHOcndOJOHHueu еносонииеи нэбас ое-иох
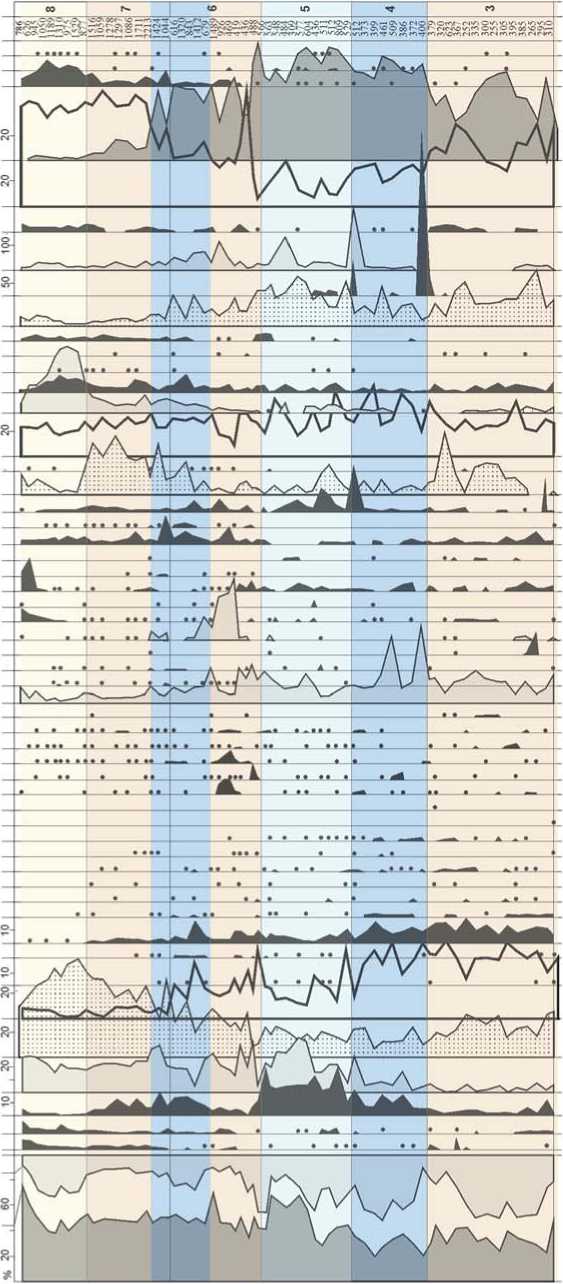
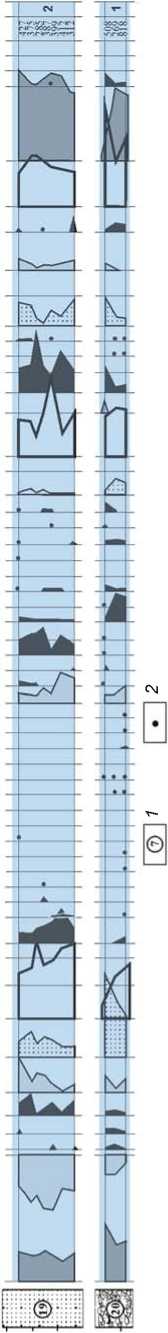
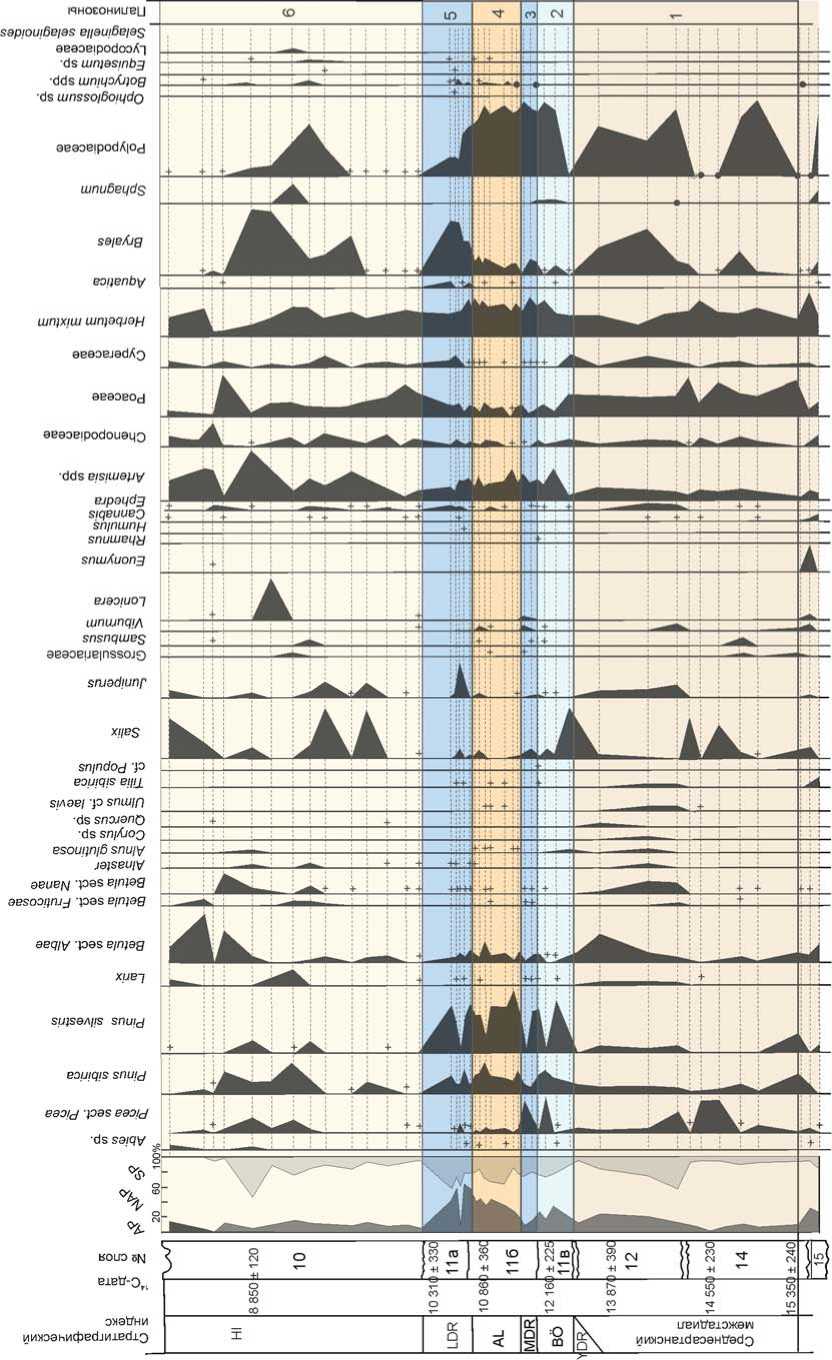
^£3888 СОСО ц*)со 6 88 БЙ СОСО КО СО ( нявебюнсеб СИчимц Я aeaoeipodouaqg аецзобшоэ etsiuiayv aeaoeiiXqdoAjeo эeJeJ!||эqшn ejoyiAjed sueqediui curuediAiA umuo6Aiod шnluoшe|Od aeau^ejg деием pas e/njag aesooiinjd pas einjag jG^seuiv auqdea eoiyeqiea зпишецу бврюишвф aeqdoddiH eaejidg jaiseauoioQ sn6aeiejQ еоипцзриеш suei6np sn/njaq snuidieo jaoy snojano ds n 'siAaei p апш/п eouiqis eii!± еиецвАв snjAjOQ snuiy aeqiv pas eppg Я эпоха ds eppg aepisoj pas e/npg suiseAjis snuid ”" eieAoqo eaoid eouiqis snuid eouiqis xueq eouiqis saiqy RdOUQ aoxhHHdeioAx и HHHaioed xiAioMHBsedi eh4ui4ij _ eoxHHdeioAx и 8эя8эdэtr еЬяшяц Я Рис. 3. Спорово-пыльцевая диаграмма нижней части плейстоценовых отложений Денисовой пещеры (аналитик Е.М. Малаева). 1 - литологический слой; 2 - содержание пыльцы менее 1%. Верхний неоплейстоцен На протяжении верхнего неоплейстоцена в окрестностях Денисовой пещеры, стоянок Усть-Каракол и Ануй-2 происходили постепенное снижение роли лесной биоты, сокращение площади лесов и расширение участков, занимаемых открытыми травянистыми и травяно-кустарничковыми сообществами [Природная среда…, 2003, с. 330–349]. В эволюции растительного покрова казанцевского межледниковья, которое традиционно сопоставляется с подстадией МИС 5е, по палиноспектрам из слоя 20 и нижней части слоя 19 в Денисовой пещере реконструированы три фазы (рис. 4). В первую фазу доминировали сосново-березовые леса с участием клена, вяза и липы, во вторую – березово-сосновые леса с примесью липы, дуба и клена, а в оптимальную фазу межледниковья в господствовавших лесах соэдификаторами были береза и сосна, в составе древостоя более заметное участие принимали ольха, ель и широколиственные породы: орех маньчжурский, граб обыкновенный, липа сибирская, вяз гладкий, дуб и лещина. Согласно климатохроностратиграфическим исследованиям Н.С. Болиховской и А.Н. Молодькова, базирующимся на результатах ЭПР- и ИК-ОСЛ-датирова-ния и палинологического анализа континентальных и морских отложений плейстоцена, большая часть стадии МИС 5 характеризуется межледниковым климатом, а возраст последнего межледниковья, имевшего по крайней мере три климатических максимума и два эндотермальных похолодания, оценивается в интервале от 145–140 до 70 тыс. л.н. [Молодьков, Боли-ховская, 2011; Molodkov, Bolikhovskaya, 2006, 2009]. Поэтому Н.С. Болиховская не исключает, что слои 19 (верхняя часть), 17 и 14 в Денисовой пещере и слои 17–14 стоянки Усть-Каракол (рис. 5), отнесенные ранее к климатической фазе, переходной к ермаковско-му похолоданию (подстадии МИС 5a – d) [Природная среда…, 2003, с. 264–270], могли формироваться во время казанцевского межледниковья, поскольку в составе палиноспектров из этих слоев отмечено довольно высокое содержание пыльцы березы, сосны обыкновенной, ольхи и широколиственных пород. К относительно холодному ермаковскому времени (МИС 4) в разрезах плейстоценовых отложений долины Ануя относятся слои 13–12 в Денисовой пещере (см. рис. 4) и на стоянке Усть-Каракол (см. рис. 5). В период формирования этих отложений отмечено три фазы развития природной обстановки. Первая фаза отражает самый неблагоприятный в климатическом отношении интервал. Согласно палинологическим показателям, в это время произошли существенное похолодание и аридизация климата, что вызвало расширение степных участков с преобладанием в травяно-кустарничковом покрове полынно-злаковых и раз- нотравно-полынно-злаковых сообществ, сокращение площади лесных стаций и уменьшение в их составе таксономического разнообразия широколиственных деревьев. Для следующей фазы, отличающейся холодным и влажным климатом, характерны отложения с высоким содержанием пыльцы хвойных пород – ели и кедра, небольшой долей пыльцы березы и отсутствием пыльцы широколиственных растений. На заключительном этапе ермаковского времени произошло общее улучшение климатической обстановки в условиях более сухого и теплого климата – заметно сократились массивы еловой тайги, восстановились площади березовых лесов с участием ольхи и лещины. Для каргинского времени (МИС 3), представленного слоем 11 в Денисовой пещере и слоями 11–8 разреза Усть-Каракол, установлены условия относительно прохладного и влажного климата. В это время в долине Ануя преобладали еловые леса с участием кедра. В оптимальные интервалы в составе лесов возрастала роль березы, появлялись вяз, липа и лещина. В разрезе стоянки Карама каргинскому времени отвечает ископаемая почва, залегающая в толще субаэральных покровных отложений. Палиноспектры из погребенной почвы (см. рис. 2) свидетельствуют о том, что в период ее формирования в окрестностях стоянки произрастали густые еловые леса, т.е. темнохвойные таежные формации в долине Ануя спускались до современной верхней границы горно-лесостепного пояса. В перигляциальных условиях сартанского ледникового периода (МИС 2) в разрезе Карамы (см. рис. 2) происходила седиментация покровных лессовидных суглинков. В это время площади лесов значительно сократились, доминирующую роль стали играть тундровые и степные ценозы. На раннем сартанском этапе среди лесных формаций преобладали разреженные сосново-лиственнично-еловые ассоциации. Значительные площади занимали криофитные кустарники Betula sect. Nanae, степные и луговые сообщества. На позднем этапе в условиях возросшей криоаридизации климата господствовали тундро-степные ландшафты. В растительном покрове были обильно представлены криофиты: Alnaster fruticosus (Duschekia fruticosa), Betula sect. Nanae, B. sect. Fruticosae, Diphazium alpinum и ксерофиты: Artemisia subgenus Dracunculus, Seriphidium. Небольшие лесные участки состояли из сосны Pinus sylvestris и кедра P. sibirica. Растительный покров сартанского похолодания в окрестностях Денисовой пещеры отражен в пали-носпектрах из слоя 9 в центральном зале пещеры, слоев 4–2 стоянки Усть-Каракол и толщи слоев 12–5 стоянки Ануй-2 (рис. 6). На протяжении большей части сартанского времени в условиях холодного и сухого климата в составе ландшафтов преобладали открытые травяно-кустарничковые и кустарниковые сообщества. Динамику эдификаторов на небольших участках Рис. 4. Спорово-пыльцевая диаграмма верхней части плейстоценовых отложений Денисовой пещеры (аналитик Е.М. Малаева). 1 - литологический слой; 2 - содержание пыльцы менее 1%; 3 - наличие спор в малочисленной группе (без подсчета процентов). вносонииви Had ас оа-иоя штебрА iunssoi6oiqdo иепэр eшшeJбo)d^^э uinui/inbe uinipueid шпщоАцод aeaoeipodXiod шnuБeцds nhqinqu hohsubioo ю % аваоЕиоцэ1Э Ahqjru hohsubido ю % UinipJV dM dBdOEJdlSV nqaBdWHCBd BhquA|j ejpdqdj septojejeo eipunj aBdOBUiBEqiunid a£ao£ja)sv шпклэ eisiiueyv uinijeo aBaoBipodouaqo dEaoEnAqdoAjBO efliiesind aeJЭ^l||эqшп Ejqji/ued suanediui eoiueqiru sui dBa3EUl6B)UE|d eqjosinBues eueinoiAv loas шпиобАр^ шпуиооу einpuediiij luruediAiA шпиобАрс^ штиош91оа aeau^ejo jgpeuiv 96U6N pase/njeg 9esooiiruj joas eppg S9qid e96Jids sn69eiejQ s9plOuшeцJ aeqdoddiH еэипцзриеш sue/бпг snojano sn/n)9q snuidiBQ J90V eouiqis eiiii dS П SIA96I Р8ПШ1П ds э еиепэле sniAroo snuiv эцоха ds epigg эерроэ pase/njeg aeqiv \oaseini9g sujs9Afis snuid eouiqis snuid eieAoqo e90«d eouiqis xueq eouiqis S9iqv AdOUQ еохкинВвюЛх и nnHaiDEd XR10HHB9Edl EhqyAU aoxHHdBioAx и aaqaadat/ EhsuAu Рис. 5. Спорово-пыльцевая диаграмма отложений стоянки Усть-Каракол (аналитик Е.М. Малаева). 1 - литологический слой; 2 - содержание пыльцы менее 1 %. Рис. 6. Спорово-пыльцевая диаграмма отложений стоянки Ануй-2 (аналитик Е.М. Малаева). 1 - литологический слой; 2 - |4С-даты; 3 - содержание пыльцы менее 1 %; 4 - наличие спор в малочисленной группе (без подсчета процентов); 5 - Rhododendron dahuricum; 6 - Hippophae rhamnoides. 1ЯН1И^ _ WMxad иихээьихекишюаиеи IRHOC aiRsahRunij _, ueiiqs eшшeJбoldAJЭ _ S9piout6ei9s enauiBeps _ шпр^пл шnssqБolцdo . uinuBeqds . obeias uinipodooAq _ шпи/die шп/podooA-] . шn^euE|eduJOO ujnpodooAj , шп^елер lunipodooAq _ шп/qoAjiog шnluqdsv . uinuijinbe ujnipu9id _ aeaoeipodA|Od s- Hada? e9-uo)i ojatngo 10 % aeao0joqoi9 BRgediOHced еЬятяи ejpgqdd _ eouiqis euejyf\j , uirueidojoeuj ujnnAqdoBAz__ suisaue) snjnqui aeaoeul6eqшn|cj eejgjionjQ aeaoBnAqdoA/eo _ ejauozjoos aeeoejaisvaMhodu 8 lUDISJIQ eejne)U9j f uaBqns annodu) eisuueyy luniptqdues uaBqns asi шэру °. aeaoeipodouaqo 2. эеаи1ше/э se/eoud eipjipuripj einiag 5Ц1шпц ejnieg snsooqnjj jepeuiv . seqid _ snonqiues snbaeiejQ . eounqspueui sueibnp snojeno sninpq snuidje^ eouiqis eipi _ siAaei p 8ПШ1П еиец9ле psnjAjoo esouqniB psnuiv 9eqiy pas eppg a- su;s9aiis snuid ^ eouiqis xueq eouiqis snuid eieAoqo еэоу eouiqis seiqy NdOUQ eo>ihHHdeioA)i и g-иинагоеd XRioHHbaedi еЪяшяи вохинс1е1зЛ)1 и дэяаабаО еЬяаяи лесной растительности характеризует развитие редколесий разного состава – елово-кедровых, еловых, сосновых, сосново-березовых с участием ели, сибирского кедра и лиственницы. В относительно благоприятные климатические интервалы заметной становилась роль сосново-березовых лесов с участием ольхи и широколиственных пород. Подробная характеристика изменений природной обстановки в заключительную стадию сартанского похолодания составлена по материалам палинологического изучения отложений в Каминной пещере (рис. 7), расположенной в среднегорной зоне долины р. Каракол, левого притока Ануя [Деревянко и др., 2000; Бо-лиховская, Маркин, 2002]. В период межстадиала, предшествовавшего раннему дриасу, 15,3–13,3 тыс. л.н. в районе пещеры доминировали горно-степные и горно-луговые ландшафты с преобладанием злаково-разнотравных сообществ. Основу древостоя ближайших лесных массивов составляли сибирский кедр, ель, пихта, сосна и береза. Широко были развиты придолинные ивняки и кустарниковые заросли из бересклета, калины, жимолости и др. Во второй половине бёллинга (12,2–12,0 тыс. л.н.) в этом районе, который относился к поясу перигляциальных лесостепей, преобладали степные участки, представленные разнотравно-злаковыми и марево-полынными сообществами. Экспозиционно или эдафи-чески эти участки сочетались на увлажненных участках с лесными массивами, а на скальных выходах и каменных россыпях – с сухими каменистыми степями. Наиболее увлажненные склоны и участки долин занимали елово-кедровые и сосновые леса с участием лиственницы и березы. В пойменных лесах заметную роль играли ивняки. В состав кустарникового яруса входили можжевельник, бузина, калина, крушина и др., на более высоких гипсометрических уровнях – можжевельник, кустарниковая береза Betula fruticosa, B. rotundifolia и др. В среднем дриасе (12,0–11,8 тыс. л.н.) в окрестностях пещеры господствовали перигляциальные степи, в которых ведущую роль играли злаково-разнотравные, полынные и кустарниковые ценозы, состоящие из Juniperus sp., Betula fruticosa, B. rotundifolia, Salix sp., Sambucus sp., Lonicera sp., Viburnum sp. и др. В долинах сохранились редкие лесные участки, представленные парковыми лиственнично-сосново-еловыми ассоциациями. Для аллерёда (11,8–10,8 тыс. л.н.) характерна наиболее благоприятная климатическая обстановка. В составе лесов постоянно участвовали липа, вяз и ольха. В термоксеротическую субстадию доминировала степная растительность с полынными и злаковоразнотравными группировками, небольшие участки лесов занимали елово-кедровые и березово-сосновые формации. В термогигротическую субстадию в соста- ве господствовавших лесостепей увеличились площади лесных массивов, возросла роль кедра, липы и вяза. Среди степных ценозов преобладали злаково-разнотравные ассоциации. В криогигротическую субстадию позднего дриа-са (10,8–10,4 тыс. л.н.) этот район занимали перигляциальные горно-лесные ландшафты. Леса были представлены ассоциациями хвойных пород – лиственницы, кедра и сосны. Широкое развитие имели кустарниковые формации из можжевельника и ольховника, кустарниковых видов березы и ивы. Встречались остепненные злаково-разнотравные и марево-полынные участки, а также слабозадернованные субстраты и каменистые ро ссыпи с Ephedra sp., Goniolimon speciosum, Cryptogramma sp. и др. На относительно суровый климат этого интервала указывает наличие пыльцы Alnaster fruticosus, Betula fruticosa, B. sect. Nanae, Botrychium boreale. В криоксеротическую субстадию позднего дриа-са (10,4–10,0 тыс. л.н.) в районе пещеры развивалась перигляциальная горная полупустыня с полынными и злаково-разнотравно-полынными сообществами, в которых заметную роль играли эфедра, маревые – Chenopodium polyspermum, Suaeda cf. physophora и представители Asteraceae, Cichoriaceae, Zygophyllaceae и др. Присутствие в спектрах пыльцы Alnaster fruticosus, B. sect. Nanae, Claytonia sp. свидетельствует о холодном и сухом климате. Согласно палинологическим данным, на заключительной стадии сартанского времени среднегорный район бассейна Ануя неоднократно находился в зоне распространения и последующей миграции степного, лесостепного и лесного поясов. Аркто-бореальные виды – Betula sect. Fruticosae, B. sect. Nanae, Alnaster fruticosus, Botrychium boreale и др. – постоянно участвовали во флоре как стадиальных, так и межстадиальных интервалов, однако тундровые, тундровостепные или тундрово-лесостепные перигляциальные формации в качестве зональных образований, скорее всего, не проникали на эту территорию. Совместное присутствие в составе автохтонных палиноспектров пыльцы Alnus glutinosa и микротермных широколиственных пород – липы сибирской Tilia sibirica и вяза гладкого Ulmus cf. laevis, не произрастающих в настоящее время в этом районе и сходных по сохранно сти с микроостатками криофитов, подтверждает выводы геоботаников о рефугиальной природе растительного покрова Северо-Западного Алтая. Заключение Детальное палинологическое изучение отложений многослойных палеолитических памятников Северо-Западного Алтая позволило реконструировать Рис. 7. Спорово-пыльцевая диаграмма плейстоценовых отложений в пещере Каминной (аналитик Н.С. Болиховская). Hl – голоцен; LDR – поздний дриас; AL – аллерёд; MDR – средний дриас; BÖ – бёллинг; YDR – древний дриас; AP – пыльца деревьев и кустарников; NAP – пыльца трав и кустарничков; SP – споры. процесс развития растительного покрова в межледниковые и холодные этапы плейстоцена, в периоды обитания здесь первобытного человека. В межледниковые эпохи нижнего неоплейстоцена в пределах низ-когорья Северо-Западного Алтая в составе степных, лесостепных и лесных ландшафтов наряду с хвойными и березовыми формациями участвовали широколиственные леса. В климатические оптимумы ниж-ненеоплейстоценовых межледниковий эта территория входила в трансконтинентальный пояс неморальных лесных и лесостепных формаций, занимавших южные районы Северной Евразии. Одна особенность алтайских неморальных лесных сообществ нижнего неоплейстоцена – совместное произрастание древесных пород, обитающих в настоящее время в далеко расположенных друг от друга центрах неморальной флоры – в районах Восточной Европы, Кавказа, Крыма и на юге Дальнего Востока. Другой важной особенностью нижненеоплейстоценовых лесов долины Ануя был состав эдификаторов – орех Juglans mandshurica, европейские и маньчжурские виды граба Carpinus betulus, C. cordata, C. orientalis и липы Tilia cordata, T. amurensis, T. mandshurica, T. sibirica. В отличие от них, в современных европейских и дальневосточных широколиственных и хвойно-широколиственных лесах основную лесообразующую роль играют различные виды дуба: Quercus robur, Q. petraea, Q. pubescens, Q. macranthera и др. – в Восточной Европе, Крыму и на Кавказе, Quercus mongolica, Q. dentata, Q. crispula и др. – на юге Дальнего Востока. В холодные эпохи нижнего неоплейстоцена в окрестностях стоянки Карама, находящейся в настоящее время в зоне перехода от горно-лесного к горнолесостепному поясу, доминировали перигляциальные степи, березовые, хвойные и хвойно-березовые редколесья и микротермные кустарниковые формации. В период сартанского похолодания здесь господствовали тундрово-степные сообщества. В современном горно-лесном поясе в районе Денисовой пещеры климатофитоценотические особенности межледниковых и холодных эпох среднего и верхнего неоплейстоцена были существенно иными. В периоды похолоданий здесь повышалась общая увлажненность и заметно расширялись площади темнохвойных еловых и кедровых лесов, ранее занимавших верхние ярусы горных склонов. Межледниковым этапам отвечали относительно сухие климатические условия и широкое распространение лесных или лесостепных зональных типов растительности, в пределах которых в составе лесных сообществ преобладали березовые и сосново-березовые древостои с заметным участием широколиственных деревьев. Развитие широколиственных лесных формаций на территории Северо-Западного Алтая в эти межледниковья не зафиксировано. Анализ спорово-пыльцевых диаграмм и составленные по ним характеристики палеорастительности свидетельствуют о том, что в межстадиальные и переходные интервалы холодных эпох в бассейне Ануя в условиях расчлененного рельефа и фрагментарного распространения многолетнемерзлых пород наряду с типично перигляциальными биотопами существовали участки с благоприятными эдафическими и мезоклиматическими условиями, на которых в пределах низкогорья произрастали березово-сосновые и березово-сосновые парковые леса с примесью широколиственных пород – дуба, липы, вяза, лещины, а в среднегорье, где расположена Каминная пещера, – елово-кедровые и березово-сосновые формации с участием липы сибирской и вяза. По составу палинофлор из средне- и верхне-неоплейстоценовых отложений прослежена тенденция уменьшения роли, а также постепенного исчезновения широколиственных пород в лесных биомах долины Ануя к началу голоцена. Установлено, что ольха черная Alnus glutinosa и широколиственные древесные породы – орех маньчжурский Juglans mandshurica, граб обыкновенный Carpinus betulus, дуб Quercus, липа сибирская Tilia sibirica, вяз гладкий Ulmus cf. laevis, клен Acer и лещина Corylus avella-na – постоянно участвовали в лесах Северо-Западного Алтая на протяжении неоплейстоцена. Реконструкции растительности Северо-Западного Алтая в межледниковые и холодные этапы неоплейстоцена указывают на то, что по степени зональной и формационной дифференциации растительного покрова ландшафты холодных интервалов не уступали ландшафтам, формировавшимся в условиях межледникового климата.