Палеоэкологические исследования, тафономия и генезис местонахождения Луговское

Автор: Лещинский С.В.
Журнал: Археология, этнография и антропология Евразии @journal-aeae-ru
Рубрика: Палеоэкология. Каменный век
Статья в выпуске: 1 (25), 2006 года.
Бесплатный доступ
Короткий адрес: https://sciup.org/14522512
IDR: 14522512 | УДК: 903
Текст статьи Палеоэкологические исследования, тафономия и генезис местонахождения Луговское
В начале XXI в. в Ханты-Мансийском автономном округе были впервые проведены палеоэкологические исследования плейстоценовой мегафауны с позиции влияния абиотической среды на крупных ископаемых млекопитающих, главным образом мамонтов. Объектом изучения стал палеофаунистический материал местонахождения Луговское**, одной из особенностей которого является абсолютное преобладание костей и зубов мамонтов над костными остатками других представителей мегафауны, и, конечно, “изюминка” Луговского – позвонок мамонта, пробитый палеолитическим орудием [Zenin et al., 2003]. Кроме того, радиоуглеродным методом по костям и зубам мамонтов и шерстистого носорога были получены “молодые” изотопные даты, до настоящего времени не характерные для центра Западной Сибири [Orlova et al., 2004].
Палеоэкологические исследования основаны на сравнительном анализе остатков ископаемых млекопитающих Луговского (~5 500 экз.) с плейстоценовой мегафауной из Волчьей Гривы (~500 экз.), Шестакова (более 700 экз.), Кочегура (более 150 экз.), Колывани (~50 экз.), Большедорохова (~200 экз.) и других местонахождений (более 300 экз.) Западно-Сибирской равнины*. Необходимо отметить, что значительная часть (более 4 000 экз.) коллекции Луговского, собранной в 1999–2004 гг., представлена довольно мелкими фрагментами, малопригодными для исследований. Поэтому детально изучено ок. 1 500 относительно целых костей и зубов мамонтов и 65 – других млекопитающих.
При сравнительном анализе максимально широко использовались опубликованные материалы по болезням, связанным с нарушением обмена веществ, особенно энзоотическим и эндемическим заболеваниям опорно-двигательной системы как современных, так и плейстоцен-голоценовых крупных млекопитающих, в т.ч. человека [Логгинов, 1890; Рохлин и др., 1934; Дамперов, 1939; Чепуров и др., 1955; Черкасова, 1954; Ковальский, 1974; Уразаев, 1978; Мащенко, Лещинский, 2001; Деревянко и др., 2003; Лещинский и др., 2003; и др.].
Тафономические особенности и генезис местонахождения
Массовость костей и зубов мамонтов на Луговском (более 95 % от общего количества костных остатков, без учета мелких фрагментов) сближает его с такими
* Изученные коллекции хранятся в Музее Природы и Человека, Томском государственном университете, Институте археологии и этнографии СО РАН и районных краеведческих музеях.
известными местонахождениями Азии, как Шестако-во, Волчья Грива и Кочегур, являющимися зверовыми палеосолонцами [Лещинский, 2001; Leshchinskiy, 2001; Деревянко и др., 2003; Лещинский и др., 2003; Leshchinskiy, Burkanova, 2003]. Остатки млекопитающих, обнаруженные в донных отложениях ручья, за исключением слоя современного/голоценового буровато-серого илистого осадка, преимущественно хорошей сохранности – выветривание нулевой или первой стадии (по: [Behrensmeyer, 1978]). Однако небольшая часть костей сильно выветрена (вторая стадия и выше), что говорит о мацерации некоторых трупов животных на открытом воздухе и/или существенном переотложении отдельных остатков. Значительно больший удельный вес наиболее выветренных и/или окатанных при транспортировке костей прямо свидетельствует об их переотложении и, вероятно, большем возрасте. Судя по геологической ситуации и результатам изотопного датирования практически всех ископаемых остатков (15 анализов), отложения слоя 2 сформированы в позднесартанское – раннеголоценовое время (~16,5–9,5 тыс. л.н.). Две 14С-даты (~18,2 и 30 тыс. л.н.), а также фрагменты костей и зубов в слое 1 явно говорят о переотложении какой-то части материала*. На некоторых костях выявлены различные стадии выветривания и следы механического разрушения – истирания (воздействие льдин?), что может свидетельствовать о частичном выносе остатков на поверхность при переотложении и/или неполном захоронении трупов, когда отдельные части туш оставались над поверхностью воды или вмещающего субстрата.
Многочисленные фрагменты ко стей и зубов, приуроченные к современным донным отложениям ручья, указывают на активное разрушение костеносных горизонтов и современный, а также раннеголоценовый значительный перенос костей, вымываемых талыми и паводковыми водами. В основных костеносных горизонтах большинство ископаемых остатков не повреждено, однако почти все они претерпели небольшой латеральный перенос еще в сартанское время. Так, при проходке выработок в 2002 г. и новых зачистках стенок раскопов 1999–2001 гг. не обнаружено костей в анатомическом положении, хотя отдельные скопления, по-видимому, представляли когда-то единые скелеты. Сохранность ископаемых остатков явно свидетельствует о быстром захоронении трупов или их частей в закисных условиях, какие были и существуют сейчас на данном участке. Наличие очень вязких (зыбучих) глинистых отложений из-за высокой степени обводненности за счет многочисленных родников, стока грунтовых, талых и паводковых вод создало здесь в конце плейстоцена природную ловушку (существует и в настоящее время), в которую, вероятно, попадали наиболее слабые и больные особи. Таким образом, не вызывает сомнений то, что захоронение происходило на месте гибели животных. Следовательно, формирование местонахождения обязано регулярному посещению млекопитающими данного участка во второй половине сартанского криохрона. По-ви-димому, причина таких посещений заключалась в существовании в долине ручья гидроморфного зверового солонца. Солонцовыми, вероятно, являлись обогащенные дефицитными макро- и микроэлементами породы цоколя надпойменных террас, а их высокая гигроскопичность способствовала образованию грязевых ванн (чем, по сути, являются донные отложения), так любимых животными-литофагами [Паничев, 1990]. Косвенным доказательством привлекательности этих пород для травоядных млекопитающих в настоящее время служат многочисленные следы присутствия лосей на подобном грязевом участке в нескольких километрах от местонахождения. Комбинация условий “зверовой солонец – ловушка” привлекала также плотоядных животных и древнего человека, поскольку охота на слабого/больного зверя, увязшего в зыбуне, вероятно, не составляла большого труда, не исключено также, что имело место трупопоедание. Обнаруженные в 2002 г. многочисленные палеолитические артефакты красноречиво говорят об этом, особенно позвонок мамонта с застрявшим кварцитовым вкладышем от метательного орудия*.
Палеоэкологические исследования
Выдвинутая гипотеза о зверово-солонцовой природе местонахождения Луговское, а также обнаруженный уникальный позвонок мамонта с пробоиной от метательного орудия явились предпосылкой для детальных палеоэкологических исследований [Павлов и др., 2002; Лещинский, в печати]. Одной из основных задач в изучении ископаемых остатков было выявление следов заболеваний опорно-двигательной системы на костях и зубах (в меньшей степени) вымерших животных.
В результате макроскопического наблюдения обнаружено ок. 780 остатков Mammuthus primigenius Blum. с деструктивными изменениями костных и соединительных тканей, 3 – Bison sp., 1 – Coelodonta antiquitatis Blum.; на костях и зубах других млекопитающих нарушения не выявлены. Таким образом, в
Относительное распределение выявленных при макроскопическом исследовании деструктивных изменений в костях различных отделов скелета мамонта с местонахождения Луговское
|
Кости и их фрагменты * |
Всего исследовано |
Выявлено с деструктивными изменениями |
Процент поражения |
|
Череп и нижняя челюсть |
~40 |
~30 |
~75 |
|
Позвонки |
~490 |
~345 |
~70 |
|
Ребра |
~250 |
~90 |
~36 |
|
Грудина |
7 |
1 |
~14 |
|
Лопатки |
26 |
13 |
50 |
|
Тазовые кости |
28 |
18 |
~64 |
|
Длинные кости |
175 |
117 |
~67 |
|
Коленные чашки |
5 |
5 |
100 |
|
Кости кисти и стопы |
~250 |
~140 |
~56 |
* Возможно, некоторые фрагменты принадлежат одной и той же кости.
коллекции Луговского содержание остатков мамонтов с признаками остеодистрофии превышает 50 % от их общего количества (мелкие фрагменты не учитывались)*.
Следует иметь в виду, что выявленный процент не отражает истинного соотношения больных и здоровых особей, т.к. часть костей в скелете животного, страдающего заболеванием опорно-двигательной системы, особенно на начальной стадии, о стается нормальной. Для получения наиболее достоверной патологоанатомической картины необходимо оцени-
* При абсолютном преобладании ископаемых остатков Mammuthus primigenius Blum. в работе рассмотрены абиотические аспекты экологии только этого вида.

Рис. 1. Проксимальный эпифиз лучевой кости (radius) M. primigenius Blum. (коллекционный № ХМ-8190/126, полевой № 137). Бороздки трения (показаны желтыми стрелками).
вать состояние различных отделов и костей скелета (см. таблицу ). Дополнительно были просмотрены зубы мамонтов и их фрагменты (более 60), из которых ~34 % имеют признаки нарушений.
Деструктивные изменения прекрасно диагностируются на эпифизарных частях костей конечностей (особенно дистальных отделов – запястных, пястных, предплюсневых, плюсневых, фалангах, а также коленных чашках), позвонках и ребрах. Наиболее распространенными и интересными являются бороздки трения и участки полного истирания компактного слоя на суставных поверхностях, возникшие в результате некроза и полного рассасывания гиалинового хряща с последующим обнажением и пришлифовкой трущихся костей (рис. 1, 2). Обычно
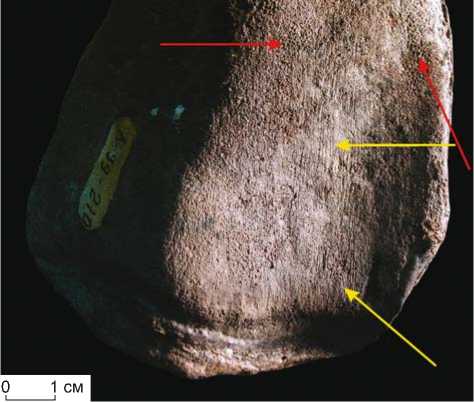
Рис. 2. Коленная чашка (patella) M. primigenius Blum. (коллекционный № ХМ-10933/39, полевой № Л-99-210). Бороздки трения (показаны желтыми стрелками), участки истирания компактного слоя (красные стрелки).

Рис. 3. Таранная кость (astragalus) M. primigenius Blum. (полевой № Л-99-230). Бороздки трения (показаны желтыми стрелками), участки истирания компактного слоя (красные стрелки), обширная язва (зеленая стрелка).
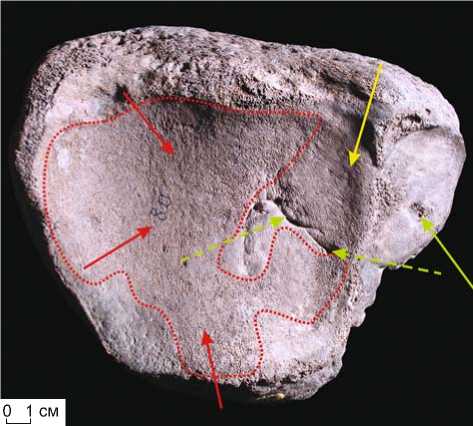
Рис. 5. Дистальный эпифиз большеберцовой ко сти (tibia) M. primigenius Blum. (полевой № 3842). Бороздки трения (показаны желтой стрелкой), зона истирания компактного слоя (красные стрелки, граница зоны – пунктирная линия), язвы (зеленые стрелки, пунктирная – частичное заживление).
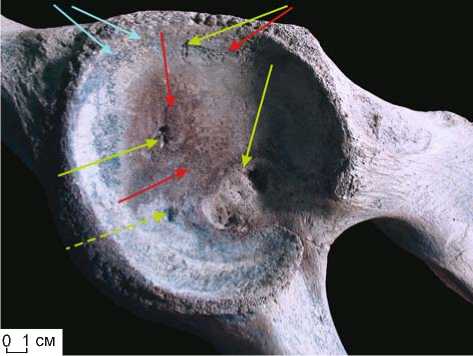
Рис. 4. Вертлужная впадина (acetabulum) таза M. primigenius Blum. (полевой № 1/93). Участки истирания компактного слоя (показаны красными стрелками), язвы (зеленые стрелки, пунктирная – частичное заживление), фиссуры (синие стрелки).

Рис. 6. Дистальный эпифиз локтевой кости (ulna) M. primigenius Blum. (полевой № 138). Выраженный остеопороз – рарефикация ко сти (расширение пор, костномозговых пространств, исчезновение компактного слоя и т.д.) с формированием грубопетлистой “сотовой” структуры.
данные изменения сопровождаются узурами, фиссурами и язвами на поверхностях суставов (рис. 3, 4). Важно отметить выявленные случаи частичного зарастания (заживления) дефектов (см. рис. 4, 5), что говорит о временном/периодическом выздоровлении животного. Широко представлены разнообразные картины системного остеопороза (истончение костных перекладин, образование пустот, расширение пор, костномозговых пространств, гаверсовых и фолькмановских каналов и др.), главным образом на эпифизах длинных костей (рис. 6, 7), позвонках (рис. 8, 9)*, ребрах и костях черепа. На позвонках и ребрах встречены “опухолевидные” разрастания костной ткани по краям суставных поверхностей, каверны и пустоты, что указывает на фиброзную остеодистрофию (и/или остеохондропатию) с образованием опухолевых масс, костных кист, остеохон-

Рис. 7. Проксимальный эпифиз лучевой кости (radius) M. primigenius Blum. (коллекционный № ХМ-8190/126, полевой № 137). Выраженный остеопороз – рарефикация ко сти (расширение пор, костномозговых пространств, исчезновение компактного слоя и т.д.) с формированием грубопетлистой “сотовой” структуры.

Рис. 8. Фрагмент эпифиза тела позвонка (corpus vertebrae) M. primigenius Blum. (полевой № 1417). Выраженный остеопороз – рарефикация ко сти с формированием мелкоячеистой структуры (образование множественных сквозных пор).
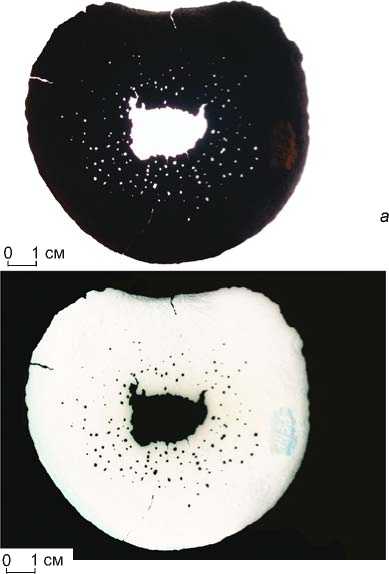
Рис. 9. Эпифиз тела позвонка (corpus vertebrae) M. primigenius Blum. (полевой № 1433). Выраженный остеопороз – рарефикация кости с формированием мелкоячеистой структуры (образование множественных сквозных пор, остеонекроз центральной зоны эпифиза).
Режим съемки – источник освещения находится за объектом:
а – позитив, б – негатив.
дром, остеоид-остеом и остеобластом (рис. 10). Для ребер также характерны искривления, периоститные явления, недоразвитость головок и их остеолиз, поперечные переломы с неполным сращением, образованием ложных суставов и/или костных мозолей. Кроме того, у трубчатых костей заметны признаки атрофии и некроза эпифизарного хряща (иногда, возможно, остеонекроза), что выражается в несра-щении эпифизов с диафизом. Аналогичные изменения часто наблюдаются на позвонках, тела которых
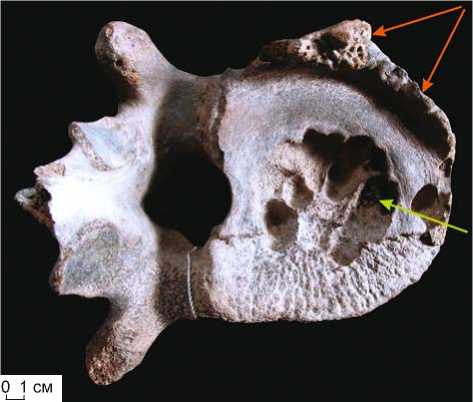
Рис. 10. Грудной позвонок (vertebra thoracica)
M. primigenius Blum. (полевой № 3691). Яркая картина остеофиброза – образование остеобластомы “сотового” типа (показана зеленой стрелкой) и периферического экзостоза (оранжевые стрелки) тела позвонка при сопутствующем остеопорозе костной ткани.
нередко имеют клиновидную форму*. Таким образом, патологический процесс разрушения хрящевой и костной тканей в той или иной мере затрагивает весь скелет. Причем поверхностные проявления заболевания в наименьшей степени заметны на грудной ко сти и ребрах (см. таблицу ).
Как показали результаты рентгеноскопии позвонка мамонта**, усиленная резорбция кости со стороны губчатой ткани, ведущая к остеопорозу, внешне может быть слабо выражена. Поэтому доля пораженных ископаемых остатков, очевидно, несколько выше, чем выявлено. Таким образом, можно говорить о практически поголовном заболевании мамонтов (не менее 90 %), остатки которых обнаружены на местонахождении. Точно определить долю здоровых животных в популяции весьма затруднительно и даже невозможно, т.к. часть мамонтов могла вовсе не посещать зверовой солонец или приходить на аналогичный объект в другой части ареала обитания. Тем не менее осмелюсь утверждать, что эта доля была ничтожно мала, а следовательно, в районе Луговского в конце сартанского криохрона имел место крупный энзоотический очаг болезни, несомненно незаразного характера, выражающейся в тотальном поражении опорно-двигательной системы мамонтов. Здесь уместно отметить, что в современных энзоотических районах, где заболеваниями поражено до 50 % и более поголовья сельскохозяйственных животных определенного вида, падеж молодняка (до 1 года) составляет 10–50 %, в тяжелых случаях даже 100 %, при средней смертности до 75 % от числа заболевших. Следовательно, в ныне существующих неблагоприятных геохимических ландшафтах часть особей адаптируется к абиотической среде обитания. Однако бесплодие самок в таких условиях достигает 50 %. Более того, в энзоотических очагах некоторые виды травоядных млекопитающих без минеральной подкормки вовсе не выживают [Чепуров и др., 1955; Плотников, 1962; Ковальский, 1974]. Таким образом, палеоэкологический анализ позволяет оценить ситуацию в Луговс-ком как безнадежную (более катастрофичную, чем в современных энзоотических очагах) для нормальной жизнедеятельности популяции, которая была обречена на вымирание.
Описанная выше патологоанатомическая картина местонахождения Луговское весьма характерна для энзоотической (эндемической) остеодистрофии – бо- лезни Кашина–Бека (уровской), – главными признаками которой являются множественные деструктивные изменения хрящевой ткани и вызванные этим деформации суставов, остановка роста и самопроизвольная ломкость костей. Современные крупнейшие очаги этого заболевания локализованы в Забайкалье, Северном Китае, Корее и на Дальнем Востоке, а также менее изученные (под другими названиями) – на Южном Урале, в Якутии, Башкортостане, Татарстане, Ульяновской и Воронежской областях, Швеции и других регионах Евразии [Дамперов, 1939; Чепуров и др., 1955; Ковальский, 1974; Уразаев, 1978; и др.]. Этиология заболевания основана на глубоком нарушении обмена веществ из-за длительного (хронического) минерального голодания, связанного с острой недостаточностью в окружающих геохимических ландшафтах, в первую очередь, Ca, Mg, P и некоторых микроэлементов, а также во многих случаях с избытком Sr.
С началом сартанского криохрона на территории Западно-Сибирской равнины начали прогрессивно развиваться кислые и особенно кислые глеевые геохимические ландшафты, характеризующиеся острой нехваткой щелочноземельных и щелочных элементов. По моему мнению, это коренным образом связано с последним подэтапом (четвертичное время) активного тектогенеза, наиболее молодое максимальное проявление которого выражается комплексом надпойменных террас позднего неоплейстоцена. Резкое, прогрессирующее увеличение скорости неотектонических поднятий Западно-Сибирской плиты фиксируется приблизительно с 50 тыс. л.н. и продолжается по настоящее время, при нескольких максимумах в сартанском криохроне и голоцене [Лукина, 1997; Трифонов, 1999; Лещинский, 2000, 2001; Лещинский, Орлова, 2004; Лещинский, Бур-канова, 2005]*. Подъем территории активизировал процессы рассоления/выщелачивания на значительной части Западно-Сибирской равнины. Таким образом, геохимические ландшафты H-класса (кислые), достигнув к началу голоцена максимума своего распространения в плейстоцене, сегодня являются определяющими на основной площади изучаемого региона, занятой тундровой, лесотундровой, таежно-лесной зонами, а также заболоченными водоразделами и поймами рек. Только в таежно-лесной зоне зафиксировано 73 % случаев недостатка Ca, Mg, Na и Co, 70 – Cu, 80 – I, 55 – Mo, 50 – B, 49 % – Zn на фоне относительного избытка Sr. В тундровых, лесотундровых и заболоченных ландшафтах дефицит жизненно необходимых элементов еще более значителен, что приводит к периодическому мине- ральному голоданию современных травоядных млекопитающих [Ковальский, 1974; Перельман, 1975; Leshchinskiy, 2001; и др.].
Изложенные факты, а также палеогеографические реконструкции сартанского криохрона не о ставляют сомнений в том, что в конце плейстоцена на севере Азии (по-видимому, и Европы) были широко развиты геохимические ландшафты, в которых крупные травоядные испытывали хронический минеральный голод. Комплексные палеонтолого-стратиграфические исследования местонахождения Луговское* позволяют утверждать, что мамонты (и другие крупные млекопитающие) данного района и, вероятно, всей Западно-Сибирской равнины в сартанское время испытывали мощный геохимический стресс [Лещинский, 2001; Leshchinskiy, 2001; Мащенко, Лещинский, 2001; Деревянко и др., 2003; Лещинский и др., 2003; Leshchinskiy, Burkanova, 2003; Зенин и др., 2004]. Отражение неблагоприятного влияния кардинально меняющейся абиотической среды мы наблюдаем на остатках мамонтов с явными признаками остеодистрофии, которую можно уверенно соотнести с болезнью Кашина–Бека.
Заключение
Результаты палеоэкологических исследований абиотических связей плейстоценовой мегафауны на территории Ханты-Мансийского автономного округа, кратко представленные в настоящей работе, позволили впервые получить данные о массовом энзоотическом заболевании поздненеоплейстоценовых мамонтов в центре Западно-Сибирской равнины. Основные выводы работы определяются следующими положениями.
-
1. Выявленные на костях мамонтов деструктивные изменения (особенно с частичным заживлением (!) дефектов) подтверждают зверово-солонцовый генезис местонахождения Луговское и ярко демонстрируют геохимический стресс мегафауны, связанный с хроническим минеральным голоданием.
-
2. Высокое содержание остатков с явными признаками остеодистрофии, характерный профиль смертности мамонтов (более 40 % – неполовозрелые особи) и другие показатели Луговского полностью тождественны характеристикам открытых ранее зверовых солонцов сартанского криохрона (Шестаково, Кочегур, Волчья Грива). Определенное сходство деструктивных изменений костей мамонтов легко выявляется в палеонтологических материалах ряда европейских местонахождений, в част
ности Севска, Костенок-1 и Краков Спадзиста Стрит (Krakov Spadzista Street) [Кузьмина, Праслов, 1992; Мащенко, 1992; Wojtal, 2001]*. По-видимому, это свидетельствует о глобальном масштабе и необратимости процессов нарушения абиотических связей между мамонтовыми популяциями и геохимической средой в конце плейстоцена.
-
3. Резкий разрыв абиотических связей, вызванный кардинальным преобразованием геохимических ландшафтов в конце плейстоцена и, как следствие, длительным минеральным голоданием, по-видимо-му, стал одной из первопричин вымирания мегафауны Северной Евразии. Результаты исследований позволяют предположить, что оно носило “лавинный” и одновременно очаговый характер.
-
4. Охота на отдельных мамонтов и других крупных травоядных, страдающих болезнями, связанными с обменом веществ, из-за хронического минерального голодания, была вполне реальна и эргономична. Однако она не могла носить массовый характер в отношении мамонтов и тем более привести к их истреблению. Резкая смена экологической ситуации, фиксируемая в сартанском криохроне и отраженная в массовой смертности крупных представителей мамонтовой фауны, возможно, стала косвенной причиной появления новых (в т.ч. вкладышевых) типов палеолитических орудий. При качественном изменении биогеоценоза, неотъемлемой частью которого являлся древний человек, экологические связи последнего также изменялись и неизбежно возникали новые культурные традиции.
