Палинологический анализ Tripolium pannonicum (Jacq.) Dobrocz. на литорали побережья Белого моря
 Dobrocz. на литорали побережья Белого моря")
Автор: Елькина Н.А., Сонина А.В.
Журнал: Принципы экологии @ecopri
Рубрика: Оригинальные исследования
Статья в выпуске: 4 (54), 2024 года.
Бесплатный доступ
Объект настоящего исследования - Tripolium pannonicum, галофит, произрастающий на западном побережье Белого моря в окрестностях пос. Кереть (Керетская губа) и пос. Растьнаволок (Онежская губа). Обобщены данные палинологических исследований за период 2014-2020 гг. Высокая степень адаптированности этого растения к условиям морских побережий делает его удобным объектом для изучения механизмов солеустойчивости, а также для палинологических исследований, как наиболее информативных о жизнеспособности популяции. Цель работы - изучение состояния пыльцы у растений T. pannonicum в разных условиях затопления на литорали. Исследование выполнено на побережье Белого моря вдоль трансект на пробных площадях в естественных границах растительности в нижней, средней, верхней литорали и на супралиторали. Всего заложено три трансекты протяженностью до 150 м. Проанализировано не менее 16000 пыльцевых зерен у 40 особей T. pannonicum на 11 пробных площадях. Качество пыльцы оценивали стандартным ацетокарминовым методом. В пробах пыльцы выявлены как нормально сформированные зерна, так и дефектные, которые объединены в семь групп. Самые низкие показатели нормально сформированной пыльцы отмечены в условиях верхней литорали Керети - 82.9 ± 3.4 % и Растьнаволока - 86.6 ± 4.3 %, а также на нижней литорали и супралиторали в Керети (77.0 ± 4.3 % и 82.7 ± 6.1 % соответственно). В ходе исследования установлено, что в средней части литорали складываются оптимальные условия для развития популяции T. pannonicum. Большее разнообразие аномальной пыльцы, отмеченное в окрестностях пос. Растьнаволок, в отличие от Керети и отсутствие значимых различий по содержанию нормально сформированной пыльцы и в Растьнаволоке, и в Керети позволяют рассматривать диапазон тератоморфной пыльцы 10-30 % как ответную реакцию вида в условиях литорали. Количество нормально сформированной пыльцы достаточное, чтобы обеспечить популяции нормальное существование за счет семенного размножения растений.
Пыльца, тератоморфы, литораль белого моря
Короткий адрес: https://sciup.org/147251111
IDR: 147251111 | УДК: 58.02 | DOI: 10.15393/j1.art.2024.15443
Palynological analysis of Tripolium pannonicum on the littoral of the White Sea coast
The object of this study is Tripolium pannonicum - a halophyte growing on the western coast of the White Sea in the vicinity of the village Keret (Keretskaya Bay) and village Rastnavolok (Onega Bay). The data of palynological studies for the period 2014-2020 are summarized. The high degree of adaptability of this plant to the conditions of the sea coast makes it a convenient object for studying the mechanisms of salt tolerance, as well as for palynological studies, as the most informative about the viability of the population. The aim of the work is to study the state of pollen of T. pannonicum plants under different flooding conditions in the littoral. The study was carried out on the coast of the White Sea along transects, in sample plots within the natural boundaries of vegetation in the lower, middle, upper littoral and supralithoral. A total of 3 transects, up to 150 m long, were laid out. At least 16,000 pollen grains from 40 plants in 11 sample plots were analyzed. Pollen quality was assessed using the standard acetocarmine method. The pollen samples contained both normally formed grains and defective ones, which were combined into 7 groups. The lowest rates of normally formed pollen were noted in the conditions of the upper littoral of the village Keret - 82.9 ± 3.4 % and the village Rastnavolok - 86.6 ± 4.3 %, as well as in the lower littoral and supralittoral in the village Keret (77.0 ± 4.3 % and 82.7 ± 6.1 %, respectively). In the course of the study it was revealed that optimal conditions for the development of the T. pannonicum population are formed in the middle part of the littoral. A greater diversity of abnormal pollen was noted in the vicinity of the village Rastnavolok, in contrast to the village Keret. This fact as well as the absence of significant differences in the content of normally formed pollen in both Rastnavolok and Keret makes it possible to consider the range of teratamorphic pollen of 10-30 % as the reaction of the species in littoral conditions. The amount of normally formed pollen is sufficient to ensure a normal existence for the population due to seed propagation of plants.
Текст научной статьи Палинологический анализ Tripolium pannonicum (Jacq.) Dobrocz. на литорали побережья Белого моря
У11ИВЕРСИТГТ®
Состояние прибрежных экосистем арктических морей находится в центре внимания ученых в связи с активным освоением арктических территорий (Olsen, 2003; Андреева и др., 2008 и др.). Большое экологическое значение в условиях изменения климата и антропогенного воздействия для стабильного развития разных биотических компонентов и систем побережий Арктики играют марши – аккумулятивные ландшафты береговой зоны с отложениями в результате приливных нагонов илистого или песчаного материала с галофитной растительностью (Голуб, Соколов, 1998). Маршевая растительность отличается рядом специфических черт в силу того, что занимает низменные береговые участки суши и заливается соленой водой во время прилива.
Белое море является частью Северного Ледовитого океана. По геоморфологии оно относится к окраинным шельфовым, внутренним континентальным водоемам. Для побережья Белого моря характерен переходный климат от субарктического морского к умеренному континентальному с продолжительной, холодной зимой (средняя температура февраля -10.5 °С) и коротким, прохладным летом (средняя температура июля 13.2 °С). Белое море на 5-6 месяцев покрывается льдом. Безморозный период длится 121 день. Годовое количество осадков ≈600 мм (Назарова, 2022).
Одним из типичных маршевых растений побережья Белого моря является галофит – астра солончаковая – Tripolium pannonicum (Jacg.) Dobrocz. (Марковская и др., 2010). Вид выделяется среди других галофитов широким спектром ответных реакций на увеличение солености, тогда как другие виды могут погибнуть от солевого стресса (Нага-левский, 2001). Для T. pannonicum отмечено снижение интенсивности фотосинтеза при высоких значениях солености, что частично связано с ограничением работы устьиц в период полного заливания растений соленой водой (Бурковская, 2008). Показана отрицательная связь между уровнем воды, относительным временем нахождения под водой и количеством закрытых соцветий у этого галофита. То есть, чем продолжительнее затопление, тем больше открытых соцветий, что, как поясняют авторы, связано с опылением под водой – гидрофилией (Марковская и др., 2015). Высокая степень адаптированно- сти T. pannonicum к условиям произрастания на морских побережьях делает его удобным объектом не только для изучения механизмов солеустойчивости (Марковская и др., 2010; Макаров и др., 2012), но и объектом для палинологических исследований, как наиболее информативных о жизнеспособности популяции.
Целью исследования являлось изучение состояния пыльцы у растений T. pannonicum в пределах западного побережья Белого моря в разных условиях затопления на литорали.
Материалы
Tripolium pannonicum из сем. Asteraceae ( Compositae ) - двулетнее травянистое растение высотой 15–80 см. Стебель полый, бороздчатый, прямостоячий, ветвистый, часто раскидистый; листья более или менее мясистые, ланцетовидные. Корзинки многочисленные, диаметром 10-15 мм, образуют густое метельчато-щитковидное соцветие (рис. 1). Обертка двухрядная, листочки обертки продолговатые. Краевые цветки – ложноязычковые, венчик краевых цветков обычно голубой, иногда розоватый, реже беловатый, до 15–18 мм длиной, 1–1.5 (2) мм шириной; трубчатые цветки диска желтые; семянки большей частью голые, 1.5–2 мм длиной. Цветет в июне - августе (Марковская и др., 2010). Произрастает по берегам рек, впадающих в Белое море в зоне ежедневного заливания морскими водами на глинистых, нередко заиленных, опесча-ненных или каменистых осушках. Предпочитает илистые осушки в устьях всех рек, является пионерным видом при их зарастании. Растение гидрофильное. Бореальный, евро-азиатский вид, типичен для побережий стран Западной Европы и Великобритании, европейской Арктики, встречается в Японии, Корее, Китае; на территории России - в европейской части, в Западной Сибири, Восточной Сибири, на Дальнем Востоке (Коро-люк, 2022).
Пыльцевые зерна у астры солончаковой сфероидальной формы, чуть сплющенные с полюсов, трехбороздноапертурные. В очертании с полюса трехлопастные. Средний диаметр пыльцевого зерна до 30 мкм. Борозды длинные, глубокопогруженные. Поры округлые. Экзина толстая, с крупношиповатой скульптурой. Шипы в основании конические, к верхушке заостренные (Punt, Hoen, 2009).

Рис. 1. Tripolium pannonicum (фото Н. А. Елькиной, Растьнаволок, 2020 г.)
Fig. 1. Tripolium pannonicum (photo by N. A. Elkina, Rastnavolok, 2020)
Исследование проводилось в пределах западного побережья Белого моря на территории Республики Карелия: в Керетской губе, окрестности с. Кереть Лоухского административного района (66°16′ с.ш., 33°33′ в.д.); в Онежской губе, окрестности пос. Растьна-волок (64°22′ с.ш., 35°93′ в.д.) Беломорского района (рис. 2). В работе обобщены данные за 6-летний период исследования, с 2014 по 2020 г.
На побережье вдоль трансект от линии уреза воды во время отлива до коренного берега были заложены пробные площадки в трех зонах литорали – нижней (НЛ), средней (СЛ), верхней (ВЛ) и на супралиторали (СуЛ) в естественных границах растительности. В зависимости от типа берега длина трансект составляла от 52 до 150 м (всего в двух районах исследования заложено 3 трансекты: в Керети – 2, в Растьнаволоке – 1). В пределах пробных площадок выполнены геоботанические описания растительного покрова (табл. 1).
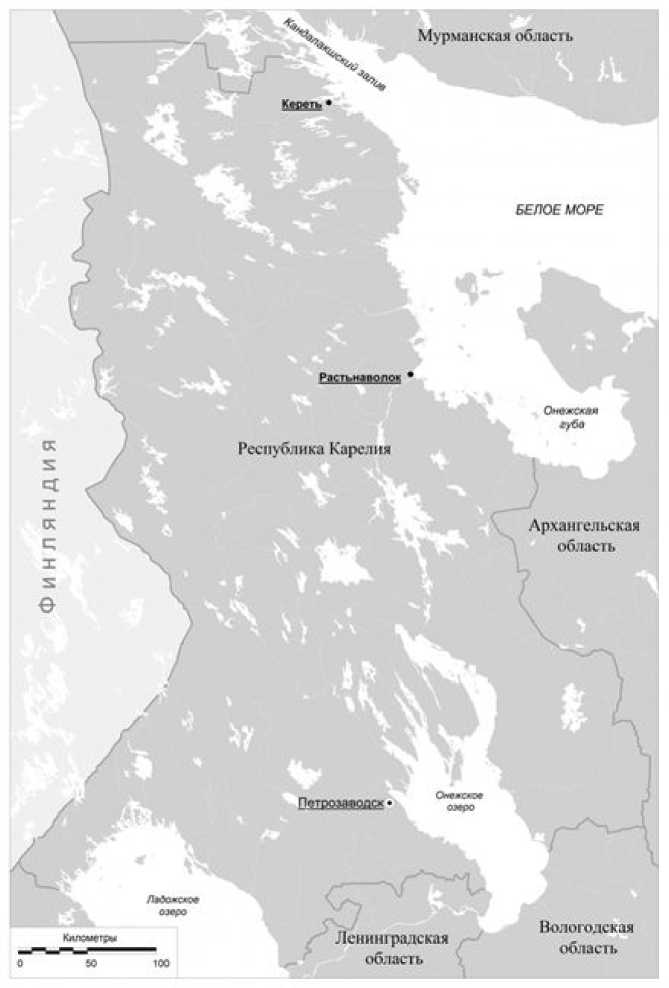
Рис. 2. Карта района исследования
Fig. 2. Map of the research area
Методы
На каждой пробной площадке были собраны соцветия с трех – четырех растущих рядом растений Т. pannonicum и зафиксированы в 70 % спирте, формируя образец данной площадки. Изучение морфологических характеристик пыльцевых зерен проведено на кафедре ботаники и физиологии растений Института биологии, экологии и агротехнологий ПетрГУ.
Оценка качества пыльцы выполнена стандартным ацетокарминовым методом (Паушева, 1980). Для приготовления ацетокармина 1 г красителя растворяли в 45 мл подогретой уксусной кислоты, добавляли
55 мл дистиллированной воды и кипятили на водяной бане в колбе с обратным холодильником в течение 1–3 часов. Краситель фильтровали горячим и хранили в посуде с притертой пробкой (Паушева, 1980). Данный метод позволяет определить и описать морфологическую структуру нормально развитых пыльцевых зерен, а также различных тератоморф. Нормально развитыми принято считать пыльцевые зерна, вегетативная клетка которых равномерно окрашена в карминово-красный цвет, с зернистой цитоплазмой, имеет вегетативное ядро и генеративную клетку с двумя спермиями. К тератоморфной (уродливой, не типичной) относят пыльцу с измененной морфологией
Таблица 1. Характеристика местообитаний растений Tripolium pannonicum на побережье Белого моря
|
Район исследования |
Общая характеристика Грунт Растительность |
|
Керетская губа |
|
|
Нижняя литораль – или- Доминирующие виды Эстуарий реки Кереть; стый грунт, средняя и на литорали: Tripolium верхняя литораль – или- pannonicum , Triglochin Окрестности пос. средняя величина при-сто-каменистый, супра- maritima , Plantago maritimа , Кереть ливов – 2.0 м, соленость , , , – 14–18 ‰ литораль – глинисто-пес- отмечено присутствие чаный и каменисто-пес- Agrostis straminea , Glaux чаный грунты maritima , Juncus atrofuscus |
|
|
Онежская губа |
|
|
Окрестности пос. Растьнаволок |
На литорали доминиру-Нижняя литораль – пес- Эстуарная зона неболь- ют виды T. maritima , P. чано-каменистый грунт, шого ручья шириной Maritima, содоминантами средняя литораль – или- 180 м; средняя величина выступают Ruppia maritima сто-песчано-каменисты , приливов – 2.0 м, соле- и T. pannonicum , отмечают- ность – 11–22 ‰ , верхняя литораль – или-ся Carex macke , nziei , Glaux сто-каменистый грунты maritima , |
(размеры, форма пыльцевых зерен, количество и тип апертур и пр.). Часто цитоплазма вегетативной клетки отходит от оболочки пыльцевого зерна и находится на разных стадиях деструкции. Пыльцевое зерно без содержимого оценивается как стерильное (Дзюба, 2007).
Для приготовления микропрепаратов из нескольких фиксированных соцветий с одной пробной площадки извлекали пыльники и помещали на предметное стекло в каплю ацетокармина, а затем механически разрушали их. Лишние ткани пыльника удаляли, препарат нагревали над спиртовкой до высыхания. Полученный препарат изучали при помощи светового микроскопа Carl Zeizz Primo Star при увеличении 400x. Для каждой пробной площади было исследовано не менее 500 пыльцевых зерен растения в трех повторностях.
Варьирование количества нормально сформированной пыльцы в пробах с разных зон литорали оценивали с помощью коэффициента вариации (СV). Уровни варьирования приняты по Г. Н. Зайцеву (1991): CV > 20% - высокий, CV...11-20 % - средний, CV < 10 % – низкий.
Результаты
Цветущие растения астры солончаковой были обнаружены в зонах нижней, средней и верхней литорали окрестностей пос. Ке-реть и Растьнаволок, а в окрестностях пос. Кереть растения произрастали и на супралиторали.
В результате палинологического анализа во всех пробах зафиксированы пыльцевые зерна с нормальной морфологией и с различными дефектами развития (табл. 2). Типичные пыльцевые зерна соответствовали описаниям в литературе (Punt, Hoen, 2009) (рис. 3) и преобладали в пробах и окрестностей пос. Кереть, и пос. Растьнаволок с незначительным уровнем варьирования в пределах литорали (CV не более 7 %) (см. табл. 2).
Все аномальные пыльцевые зерна были объединены в семь морфотипов: с измененной формой, без элементов структуры экзины, смятые пыльцевые зерна, карликовые, гигантские, с комковатой цитоплазмой и стерильные. У растений с побережья окрестностей пос. Растьнаволок зафиксирована пыльца шести патологических морфотипов, а в окрестностях пос. Кереть – трех. И в Ке-рети, и в Растьнаволоке встречается общая тератоморфа – пыльцевое зерно с нарушениями скульптуры экзины, а также стерильная пыльца.
Наиболее часто (10.8–19.6 %) стерильная пыльца (рис. 4) обнаруживается на литорали в пос. Кереть: экзина развита правильно, пыльцевые зерна нормального размера, но слабо окрашиваются или не окрашиваются кармином. В окрестностях пос. Растьнаво-лок стерильная пыльца встречается существенно реже (0.5–3.0 %). Максимальное содержание такой пыльцы и в Керети, и в Растьнаволоке выявлено в пробах для зоны нижней литорали, а в зоне средней литорали – минимальное (см. табл. 2).
Таблица 2. Качество пыльцы астры солончаковой на исследованных участках
Варианты морфологического строения пыльцы (морфотипы):
без элементов карлико- стериль- смятые гигантская комковатая скульптуры M ± S (%) вая M ± S M ± S (%) ЦП M ± S (%) ная M ± S экзины M± (%) (%)
S (%)
измененная форма M± S (%)
Зона морфоло-побе- гическая CV, режья норма M % ± S (%)
Кереть
|
НЛ |
77.0 ± 4.3 |
5 |
- |
1.5 ± 0.9 |
- |
- |
- |
1.8 ± 0.6 |
19.6 ± 3.5 |
|
СЛ |
86.3 ± 3.1 |
4 |
- |
1.7 ± 1.0 |
- |
- |
- |
1.7 ± 1.0 |
10.8 ± 2.8 |
|
ВЛ |
82.9 ± 3.4 |
4 |
- |
2.0 ± 0.8 |
- |
- |
- |
1.9 ± 0.9 |
12.7 ± 2.7 |
|
СуЛ |
81.7 ± 1.3 |
2 |
- |
1.5 ± 0.5 |
- |
- |
- |
1.0 ± 0.7 |
15.9 ± 1.4 |
|
Растьнаволок |
|||||||||
|
НЛ |
82.7 ± 6.1 |
7 |
1.9 ± 1.0 |
0.9 ± 0.7 |
0.9 ± 0.5 |
9.8 ± 3.4 |
0.7 ± 0.4 |
- |
3.0 ± 0.4 |
|
СЛ |
89.7 ± 1.2 |
1 |
3.2 ± 0.4 |
0.7 ± 0.2 |
0.5 ± 0.2 |
3.6 ± 1.7 |
0.7 ± 0.2 |
- |
0.5 ± 0.5 |
|
ВЛ |
86.6 ± 4.3 |
5 |
1.4 ± 0.4 |
5.4 ± 4.9 |
0.7 ± 0.6 |
3.8 ± 2.9 |
1.2 ± 0.9 |
- |
0.5 ± 0.5 |
Примечание. НЛ - нижняя литораль, СЛ - средняя литораль, ВЛ - верхняя литораль, СуЛ - супралитораль; CV – коэффициент вариации (%) для нормально сформированной пыльцы, ЦП – цитоплазма; M ± S – среднее арифметическое ± стандартное отклонение.
Реже встречаются тератоморфы без элементов скульптуры экзины. Пыльца стандартных размеров, хорошо окрашена кармином. Однако поверхность таких пыльцевых зерен практически гладкая, шипы существенно сглажены или отсутствуют (рис. 5). На трансекте в Керети количество пыльцы данного морфотипа составило 1.5–2.0 %, в Растьнаволоке – 0.7–5.4 %. Максимальное количество зафиксировано для зоны верхней литорали (см. табл. 2).
Только в пробах пыльцы из Керети найдены пыльцевые зерна с комковатой цитоплазмой. Пыльца имеет нормальные размеры, но цитоплазма вегетативной клетки пыльцевого зерна отходит от его стенок и неравномерно прокрашена кармином, что придает ей вид комочков. Количество такой пыльцы незначительно (1.0-1.9 %) и достигает максимума в зоне верхней литорали (см. табл. 2).
В пробах пыльцы с трансекты в пос. Расть-наволок обнаружены пыльцевые зерна нестандартных размеров. Больше всего зафиксировано карликовых пыльцевых зерен (размеры в два и более раза меньше нормы), причем других аномалий развития такая пыльца не имела (3.6–9.8 %, максимум в зоне нижней литорали). Реже встречалась пыльца, имеющая отличную от стандартной форму - вытянутые или сплющенные пыльцевые зерна (1.4–3.2 %, максимум в зоне средней литорали). Были зафиксированы смятые пыльцевые зерна (0.5-0.9 %, максимум в зоне нижней литорали). Самым редким дефектом строения в пробах пыльцы из Растьнаволока были гигантские пыльцевые зерна (в два и более раза больше нормативных) – 0.7–1.2 %, с максимумом в зоне верхней литорали. Других отклонений в развитии у них выявлено не было.
Сравнение результатов палинологического анализа показало, что количество нормально сформированной пыльцы сопоставимо в окрестностях Керети и Растьнаво-лока, отсутствие различий подтверждает и дисперсионный анализ (табл. 3).
Меньшее количество нормально развитой пыльцы выявлено в пробах из зоны нижней литорали (Кереть – 77.0 ± 4.3 %, CV 5 %; Растьнаволок – 82.7 ± 6.1 %, CV 7 %). Коэффициент вариации для нормально развитой пыльцы в зоне нижней литорали также выше (Кереть – 7 %, Растьнаволок – 5 %), чем для других зон, но не превышает 10 %. В зоне нижней литорали зафиксированы максимальные значения для большинства патологических морфотипов пыльцы.
Для зоны средней литорали характерно самое высокое содержание нормально развитой пыльцы как в Керети, так и в Растьна-волоке (86.3 ± 3.1 %, CV 4 % и 89.7 ± 1.2 %, CV 1 % соответственно). Пыльца с разными вариантами тератоморф в этой зоне литорали встречается в минимальных или средних количествах.

Рис. 3. Пыльцевое зерно Tripolium pannonicum нормального строения Fig. 3. Pollen grain of Tripolium pannonicum of normal structure
Рис. 4. Пыльцевое зерно Tripolium pannonicum без содержимого
Fig. 4. Pollen grain of Tripolium pannonicum without contents
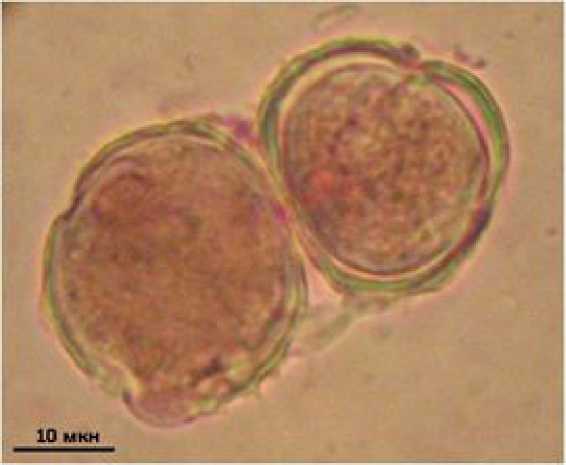
Рис. 5. Пыльцевые зерна Tripolium pannonicum без шипов экзины
Fig. 5. Pollen grains of Tripolium pannonicum without exine spines
В пробах пыльцы с верхней литорали зафиксировано среднее количество нормально сформированной пыльцы (Ке-реть – 82.9 ± 3.4 %, CV 2 %; Растьнаволок – 86.6 ± 4.3 %, CV 5 %). Тератоморфная пыльца встречается в средних или максимальных количествах.
Дисперсионный анализ выявил значимые различия по содержанию нормально сформированной пыльцы между верхней и нижней, а также между средней и нижней литоралью в условиях побережья Керети. Растения верхней и средней литорали не различаются по содержданию нормально сформированной пыльцы. Несколько иная картина выявлена для побережья Растьна-волока, где выявлены значимые различия между нижней и средней, средней и верхней литоралью (см. табл. 3). Обсуждение
T. pannonicum относится к растениям лито-
Таблица 3. Дисперсионный анализ сравнения нормально сформированной пыльцы в разных районах исследования
|
Место положения |
ВЛ – НЛ |
НЛ – СЛ |
СЛ – ВЛ |
|
Кереть |
0.0002*** |
0.001*** |
0.07 |
|
Растьнаволок |
0.4 |
0.03* |
0.04* |
|
НЛ |
СЛ |
ВЛ |
|
|
Кереть – Растьнаволок |
0.08 |
0.06 |
0.5 |
Примечание. *** – р ≤ 0.001; ** – р ≤ 0.01; * – р ≤ 0.05.
рального комплекса, хорошо адаптированным к условиям периодического заливания морской водой (Гуляева, Марковская, 2019). Выполненный пыльцевой анализ у растений двух районов исследования в пределах побережья Белого моря показал довольно стабильное состояние репродуктивной сферы, о чем свидетельствуют низкие значения коэффициента вариации нормально сформированной пыльцы (CV не превышает 7 %) (Зайцев, 1991). Во всех зонах литорали в пробах пыльцевых зерен отмечено более 70 % нормально сформированной пыльцы без нарушений морфологического строения (см. табл. 2). Тем не менее обращают на себя внимание самые низкие показатели нормально сформированной пыльцы в условиях верхней литорали двух районов исследования (в Керети – 82.9 ± 3.4 %, в Растьнаво-локе – 86.6 ± 4.3 %), а также нижней литорали и супралиторали в Керети (77.0 ± 4.3 % и 82.7 ± 6.1 % соответственно).
Зона нижней литорали характеризуется самым длительным по времени периодом заливания: в эту зону вода приходит раньше всего во время прилива и позже уходит во время отлива. Зона же верхней литорали характеризуется меньшей по времени длительностью заливания, при этом погружены в воду во время периодических приливов только нижние части растения (Гуляева, Марковская, 2019). В то время как в условиях средней литорали, для которой характерна средняя продолжительность заливания растений и большая волновая динамика, отмечаются самые высокие значения нормально сформированной пыльцы (см. табл. 2). Таким образом, продолжительность заливания оказывает влияние на качество пыльцы: в условиях более продолжительного или короткого времени заливания растений морской водой обнаружено большее число дефектной пыльцы.
Встает вопрос о том, как расценивать количественные показатели дефектности пыльцы. На основании анализа пыльцы бо- лее чем у 300 видов растений достоверно разработан критерий оценки понятия «дефектная пыльца» (Куприянов, Жолобова, 1975). Авторы считают, что содержание в зрелых пыльниках свыше 11 % дефектных пыльцевых зерен приводит к снижению семенной продуктивности, они расценивают такие растения как склонные к апомиктическому размножению. В настоящем исследовании дефектная пыльца в пробах составляет около 10 % только в условиях средней литорали окрестностей пос. Растьнаволок. На всех остальных участках содержание дефектной пыльцы превышает пороговое установленное значение, что может указывать на трудности семенного воспроизведения растений астры солончаковой практически на всех участках литорали.
В данном исследовании описано семь типов тератоморф у T. pannonicum . При этом в пробах пыльцы в Керети зафиксировано только три: пыльца с комковатой цитоплазмой вегетативной клетки, пыльца с отсутствием шипов на поверхности экзины и стерильная пыльца. Меньшее количество тера-томорфной пыльцы обнаружено у растений из зоны средней литорали. В Растьнаволоке выявлено пять типов тератоморф, четыре из которых встречены только здесь: измененной формы, смятые, карликовые, гигантских размеров.
Известно, что дефекты развития цитоплазмы чаще всего возникают вследствие нарушения процесса мейоза (Дзюба, 2007). Вероятно, комковатая цитоплазма – это начальная стадия деструктивных изменений цитоплазмы вегетативной клетки пыльцы, приводящих к полному лизису содержимого пыльцевого зерна. На строение спородер-мы наиболее сильное влияние оказывают внешние факторы среды, нарушения в развитии экзины происходят на более поздних этапах формирования пыльцевого зерна, чем деградация цитоплазмы (Дзюба, 2007). То есть, даже если изначально сформировавшиеся пыльцевые зерна не имеют мор- фологических нарушений строения, есть вероятность, что могут появляться дефекты на поздних стадиях развития пыльцы. В настоящем исследовании изменения в строении экзины отмечены в пределах всей литорали, но больше таких тератоморф обнаружено в окрестностях пос. Растьнаволок в условиях верхней литорали.
Карликовая пыльца образуется вследствие нарушений на этапе редукционного деления мейоза (Зеленцов, Мошненко, 2004). Гаметопатогенные соединения и факторы природного происхождения влияют на морфологическое развитие пыльцы, что, как правило, сказывается на способности пыльцы к оплодотворению. Гигантская пыльца формируется в связи с нарушением мейоза, в частности с нерасхождением хромосом, что в дальнейшем, после оплодотворения, приводит к формированию полиплоидов, характерных для представителей сем. Asteraceae (Barker et al., 2016), которые могут быть более устойчивыми к внешним воздействиям среды, более пластичными.
Известно также, что нарушение процессов нормального развития пыльцевых зерен возникает как ответ растительного организма на внешние воздействия (неблагоприятные факторы физической среды, антропогенные фактор) (Дубровная, Бавол, 2011; Ikeuchi et al., 2015; Ашапкин и др., 2016) и на факторы биологической природы (способ размножения, генетические факторы, например наличие хромосомных рас, неустойчивость кариотипа, внутривидовая гибридизация, цитоплазматическая мужская стерильность, ядерная мужская стерильность) (Круглова и др., 2000). В настоящем исследовании отмечена реакция репродуктивных структур на условия обитания растений. В пределах зон литорали выявлены различия по содержанию в пробах нормальной и тератоморфной пыльцы. В Керети зоны литорали довольно сближены из-за особенностей береговой линии: нижняя литораль более продолжительное время затапливается, верхняя менее подвержена затоплению. Зоны в отношении этого фактора последовательно сменяют друг друга. По содержанию нормально сформированной пыльцы не отличаются пробы из верхней и средней литорали. В условиях Растьнаволока не выявлены различия у растений верхней и нижней литорали (см. табл. 3) по содержанию нормально сформированной пыльцы, казалось бы, двух разных по продолжительности и глубине затопления участков. В силу геоморфологии береговой зоны у урезы воды находится скальный вал, который приподнимается над грунтом, где складываются условия, по продолжительности затопления близкие к верхней литорали.
На основании анатомических, физиологических и сравнительных палинологических исследований установлено, что растения T. pannonicum хорошо адаптированы к условиям литорали (Елькина, Карпова, 2015; Гуляева, Марковская, 2019). Пороговый и превышающий порог уровень дефектной пыльцы, по данным некоторых авторов (Куприянов, Жолобова, 1975), должен указывать на слабое семенное воспроизведение этого вида в данных условиях среды. Однако в результате полевых исследований можно констатировать, что вид успешно развивается. Вероятно, T. pannonicum реализует именно семенное размножение в силу того, что является одно-двулетним растением, монокарпиком. Наличие пыльцевых зерен с разной структурой экзины может обеспечивать перекрестное опыление растениям T. pannonicum и в условиях, когда растение покрывается водой (гладкая экзина) – гидрофилия и когда оно произрастает в условиях частичного затопления, без погружения в воду соцветий (экзина с шипами) – энтомофилия (Жизнь растений, 1981, 1982; Римская-Корсакова, 2001). В данном случае наличие пыльцы без шипов целесообразно рассматривать как вариант адаптированности к условиям существования на литорали.
Заключение
Выполненное исследование пыльцы Tripolium pannonicum на литорали Белого моря показало, что в средней части литорали складываются оптимальные условия для развития популяции. Большее разнообразие аномальной пыльцы, отмеченное в окрестностях пос. Растьнаволок, в отличие от Ке-рети и отсутствие значимых различий по содержанию нормально сформированной пыльцы и в Растьнаволоке, и в Керети позволяют предположить, что диапазон тера-таморфной пыльцы 10–30 % можно рассматривать как ответную реакцию T. pannonicum в условиях литорали. Количество нормально сформированной пыльцы достаточное, чтобы обеспечить популяции нормальное существование за счет семенного размножения.
