Паспортизация новых линий и гибридов подсолнечника селекции ВНИИМК с помощью биохимических и молекулярных маркеров

Автор: Гучетль С.З., Челюстникова Т.А., Антонова Т.С.
Рубрика: Селекция и семеноводство сельскохозяйственных растений
Статья в выпуске: 3 (163), 2015 года.
Бесплатный доступ
Паспортизация новых перспективных линий и гибридов подсолнечника по изоферментным маркерам и молекулярным микросателлитным локусам ДНК является важным инструментом для защиты прав селекционеров. Целью данной работы была паспортизация 12 линий и четырех гибридов подсолнечника селекции ВНИИМК. Для характеристики селекционного материала использовали методы анализа изоферментов и полимеразной цепной реакции амплификации ДНК. С использованием пяти изоферментных и восьми SSR-локусов были составлены биохимические и молекулярно-генетические паспорта. По сравнению с группами линий ВНИИМК, изученными в предыдущие годы, уникальность набора новых линий, выявляемая методом анализа вариабельности изоферментных систем, стала меньше. Анализ биохимических характеристик 12 линий показал, что они группируются в восемь групп, отличающихся друг от друга. Гибридные комбинации обладали уникальными изоферментными спектрами. Уникальность линий и гибридов, полученная при помощи анализа изоферментных систем, составила 75 %. При анализе SSR-локусов выявлена индивидуальность аллельного состава каждой линии и гибрида, за исключением линий ВКІ-ими А и ВКІ-ими Б, которые являются стерильным и фертильным аналогами. Уникальность линий и гибридов, полученная при помощи микросател- литных локусов, составила 93 %. Установлено, что полиморфными и информативными для данной группы образцов являются четыре изоферментные системы: эстераза, малатдегидрогеназа, 6-фосфо-глюконатдегидрогеназа, глюкозофосфа- тизомераза, и пять микросателлитных локусов: ORS509, ORS 595, ORS1144, ORS1796, ORS 1036. Среднее число аллелей на локус для них составило 2.
Подсолнечник, паспортизация, ssr-маркеры, изоферментные локусы
Короткий адрес: https://sciup.org/142171302
IDR: 142171302 | УДК: 631.523:633.854.78
Certification of new sunflower lines and hybrids of VNIIMK breeding by means of biochemical and molecular markers
Certification of new promising sunflower lines and hybrids on isozyme markers and molecular microsatellite loci is an important method of breeders’ rights protection. The purpose of this work was to certify 12 Unes and four hybrids of sunflower of VNIIMK breeding. To characterize the breeding material there were used methods of isozyme analysis and polymerase chain reaction of DNA amplification. Using five isozyme and eight SSR-loci, biochemical and molecular-genetic passports were done. A set of the new lines revealed by analysis of variability of the isozyme systems became a less unique in comparison to sunflower Unes of VNIIMK breeding studied in previous years. An analysis of biochemical characteristics of 12 lines showed that they can be united into eight groups differed from one another. The hybrid combinations possessed unique isozyme spectra. Singularity of the lines and hybrids showed by analysis of isozyme systems was 75%. At analyzing of SSR- loci an individuality of allele set for each line and hybrid was determined, except lines VKl-imi A and VKl-imi В being the sterile and fertile analogs. A singularity of the lines and hybrids obtained by means of microsatelUte loci was 93%. The polymorphic and informative systems for this group of samples are determined to be four isozyme systems: esterase, malatdehydrogenase, 6-phosporus-gluconatdehydro- genase, glucosephosphatisomerase, and five microsatelUte loci: ORS509, ORS 595, ORS 1144, ORS 1796, and ORS 1036. An average number of aUeles for locus for them were 2.
Текст научной статьи Паспортизация новых линий и гибридов подсолнечника селекции ВНИИМК с помощью биохимических и молекулярных маркеров
Введение. Паспортизация и идентификация линий, гибридов и сортов сельскохозяйственных культур с помощью биохимических и молекулярных маркеров широко применяется в селекционной практике. Для этих целей в качестве генетических маркеров используются и морфологические признаки. Однако количество информативных маркеров такого типа ограничено. К тому же фенотипические признаки могут иметь сложный характер наследования и часто зависят от условий внешней среды.
Биохимические и молекулярные маркеры не зависят от условий окружающей среды, являются многочисленными, в большинстве своем кодоминантно наследуются и их анализ осуществляется в лабораторных условиях в краткие сроки. С помощью биохимических и молекулярных маркеров в нашей стране и за рубежом производится генетическая паспортизация таких культур, как пшеница [1; 2], соя [3], яблоня [4], рис [5], виноград [6], подсолнечник [7; 8; 9]. Так, во Франции с использованием пяти SSR-маркеров (simple sequence repeat) паспортизировано 286 сортов картофеля с созданием базы данных, заключающей в себе аллельный состав каждого сорта [10]. Семьдесят восемь SSR-маркеров были отобраны и использованы для оценки генетической изменчивости среди набора 124 инбред-ных линий подсолнечника [11]. Согласно исследованиям Imerovski с соавторами, ДНК-локусы, ассоциированные с селекционно-ценными признаками, также являются эффективными для идентификации генотипов подсолнечника [8].
Паспортизация образцов, линий и гибридов подсолнечника селекции ВНИИМК при помощи изоферментных систем, RAPD и SSR-производных амплифициро-ванных фрагментов ДНК проводится с середины 90-х годов прошлого века [12; 13; 14; 15].
Целью данной работы была характеристика районированных и перспективных линий и гибридов подсолнечника селек- ции ВНИИМК по биохимическим – изо-ферментным маркерам и молекулярным – микросателлитным локусам ДНК.
Материалы и методы. Материалом для исследования служили 12 линий и четыре гибрида коллекции подсолнечника ВНИИМК (табл. 1, 2). Для определения однородности генотипов отбирали по 3–5 индивидуальных растений каждого образца.
Изоферменты экстрагировали гомогенизацией части семядоли сухой семянки в 20 мкл 0,1 М трис-HCl буфера с добавлением 0,1 %-ного поливинилпирролидона. Электрофорез выполняли в 12 %-ном крахмальном геле с добавлением 2 % сахарозы. Для электрофоретического разделения изоферментов использовали две буферные системы, составленные по методикам, предложенным Cardy et аl. [16]. После электрофореза гелевая пластина разрезалась тонкой нихромовой нитью на 5–6 срезов. Выявление зон энзиматической активности проводилось окрашиванием по стандартным прописям Vallejos [17]. Исследовали пять изоферментных систем, описанных ранее в работе Туркав с соавторами [15].
Для проведения ПЦР (полимеразной цепной реакции амплификации) геномную ДНК выделяли из семядольных листьев 5–7-днев-ных этиолированных проростков подсолнечника с помощью модифицированного метода Saghai-Maroof et аl. [18] с использованием СТАВ буфера. Для проведения ПЦР использовали 25 мкл реакционной смеси следующего состава: 67 мМ трис-HCl, рН 8,8; 16,6 мM сульфата аммония; 1,5–3 мM MgCl2; 0,01 % Tween 20; по 0,2 мM дезоксирибонуклеозидфосфатов; по 10 пМ праймеров; 10 нг матричной ДНК и 1 ед. рекомбинантной термостабильной ДНК полимеразы (Сибэнзим, Москва). Для амплификации применяли термоциклер S1000тм (BioRad, США). Условия амплификации: начальная денатурация при 96 оС в течение 2 мин; затем 30 циклов при соблюдении температурно-временного режима: отжиг при 60 оС в течение 40 с, элонгация – 1 мин при 70 оС, денатурация при 94 оС – 30 с, финальная элонгация – 2 мин. Использовали восемь SSR-локусов, наследование которых было установлено исследованиями Челюстниковой [14], Гу-четль с соавторами [13] и Antonova с соавторами [19].
Электрофорез продуктов амплификации проводили в агарозном геле (2 % агароза, 1 х ТАЕ-буфер) с использованием камеры для горизонтального электрофореза (SE.2, ДНК-технология, Россия) в течение 1–1,5 часов при силе тока 58 mA и напряжении 90–100 V. Последующее окрашивание осуществляли бромистым этидием. Визуализация результатов электрофореза в ультрафиолетовом свете и их документирование обеспечивались при помощи системы цифровой документации видеоизображения BIO-PRINT (Vilber Lourmat, Франция).
Результаты и обсуждение. По результатам предыдущих работ для характеристики линий и гибридов по биохимическим маркерам были выбраны полиморфные изоферментные системы с примерно равной частотой встречаемости аллозимных вариантов [12; 15]. Это эстераза (EST), малатдегидрогеназа (МDH), 6-фосфоглюконатдегидрогеназа (6PGD), глюкозофосфатизомераза (GPI), глутаматдегидрогеназа (GDH). На рисунке 1 представлены энзиматические электрофоретические спектры некоторых изученных образцов.
А.

Б.
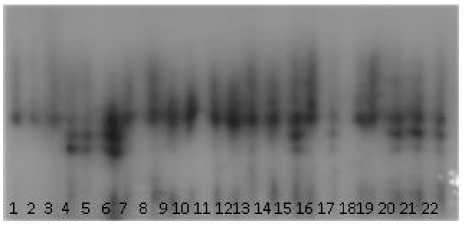
В.
■

12345678 910111213141516171819 20 2122
Рисунок 1 – Электрофоретические спектры изоферментных систем подсолнечника:
А – глюкозофосфатизомераза (GPI);
Б – малатдегидрогеназа (MDH);
В – эстераза (EST).
Расположение на дорожках: 1–3 – ВК 302,
4–6 – ВК 101, 7–9 – RG, 10–12 – Р 96, 13–14 – ВК 551(К), 15–19 – Арсенал, 20–22 – Фактор
Изоферментные спектры контролируются локусами с 2–3 аллелями, которые обозначены как: SS – контролирующий аллозим с наиболее медленной электрофоретической подвижностью, FF – алло-зим с быстрой электрофоретической подвижностью, vFvF – аллозим с наибольшей электрофоретической подвижностью. Обычно гомозиготные генотипы представлены на фореграммах одной фракцией. Исключение составляют образцы подсолнечника с генотипом Mdh – SS, представленные тремя фракциями. Гетерозиготные генотипы, в зависимости от четвертичной структуры фермента, представлены 2–5 фракциями.
Образец считали однородным, если все проанализированные семянки имели идентичный генотип. В случае если образец был неоднородным, то за типичный генотип принимали преобладающий. По всем инбредным линиям и гибридам были составлены изоферментные паспорта (табл. 1, 2).
Количество аллелей для различных локусов варьировало от 1 до 2. Среднее число аллелей на локус составило 1,6 (с учетом мономорфных локусов) и 2 (без учета мономорфных локусов). Анализ биохимических характеристик 12 линий показал, что они группируются в восемь групп, отличающихся друг от друга (табл.
Таблица 1
Изоферментные фенотипы 12 инбредных линий подсолнечника
ВНИИМК, г. Краснодар, 2015 г.
|
Линия |
Изоферментный фенотип |
||||
|
EST |
MDH |
GPI |
PGD |
GDH |
|
|
ВК 1ими А |
FF |
SS |
FF |
SS |
SS |
|
ВК 1ими Б |
FF |
SS |
FF |
SS |
SS |
|
BK 22 ими |
FF |
SS |
FF |
SS |
SS |
|
BK 301 |
FF |
SS |
FF |
FF |
SS |
|
ВК 930 |
FF |
SS |
FF |
FF |
SS |
|
ВА 760 |
vFvF |
FF |
SS |
FF |
SS |
|
P 96 |
FF |
FF |
SS |
FF |
SS |
|
ВК 302 |
FF |
FF |
FF |
FF |
SS |
|
ВК 101 |
SS |
SS |
SS |
FF |
SS |
|
RG |
SS |
FF |
FF |
FF |
SS |
|
ВК 876 |
SS |
SS |
FF |
SS |
SS |
|
ВК 195 |
SS |
SS |
FF |
SS |
SS |
Таким образом, в некоторые группы попало несколько линий, имеющих сходный генотип по составу изоферментов. Уникальность анализируемой коллекции линий составила 67 %. Столь низкий процент уникальности вызван отчасти тем, что линии ВК 1 ими А и ВК 1 ими Б являются стерильным и фертильным аналогами соответственно, и должны совпадать по всем остальным признакам.
Линии ВК 876 и ВК 195 также обладают сходным спектром изоферментов. Тем не менее, необходимо признать, что по сравнению с группами других линий ВНИИМК, изученных нами в предыдущие годы также методом анализа вариабельности изоферментных систем, процент уникальности новых линий становится меньше. Так, уникальность анализированных по изоферментным локусам 18 селекционных инбредных линий подсолнечника коллекции ВНИИМК в 1996 г. составила 78 % [15]. Уникальность 32 линий подсолнечника из той же коллекции, анализированной в 2004 г., составила 75 % [12]. Кроме того, некоторые изоферментные локусы, ранее характеризовавшиеся наличием нескольких аллелей, при изучении новых линий показывают только одно аллельное состояние. В более ранних исследованиях Туркав с соавторами [15] и Гучетль с соавторами
[12] паспортизация селекционного материала выполнялась по полиморфным системам, в состав которых входили эстераза, малатдегидрогеназа, 6-фосфоглюко-натдегидрогеназа, глюкозофосфатизомераза, глутаматдегидрогеназа. У изученно-го в 2015 г. набора линий, полиморфизм по глутаматдегидрогеназе выявлен не был. Уменьшение полиморфизма селекционного материала могло произойти либо из-за происхождения ин-бредных линий из одного или небольшого числа источников, либо из-за различной адап-тивной ценности аллельных вариантов, приводящей к преобладанию в коллекции одного из аллелей.
Четыре исследованных гибрида имели отличающиеся изоферментные спектры (табл. 2). Совокупный процент уникальности анализированных линий и гибридов составил 75.
Таблица 2
Изоферментные фенотипы четырех гибридов подсолнечника
ВНИИМК, г. Краснодар, 2015 г.
|
Гибрид |
Изоферментный фенотип |
||||
|
EST |
MDH |
GPI |
PGD |
GDH |
|
|
Имидж |
FF |
SS |
FF |
SS |
SS |
|
Арсенал |
FvF |
FS |
FS |
FF |
SS |
|
Фактор |
FS |
FS |
FS |
FF |
SS |
|
Окси |
SS |
SS |
FF |
SS |
SS |
Гибриды Фактор и Арсенал гетерозиготны по трем изоферментным локусам – Est, Mdh, Gpi . В силу того, что родительские линии гибридов Имидж (ВК 1 ими и ВК 22 ими) и Окси (ВК 876 и ВК 195) имели идентичные фенотипы, у этих гибридов нет изоферментных локусов в гетерозиготном состоянии. Следовательно, определение уровня гибридности по изо-ферментным локусам невозможно. Но возможно определение генетической чистоты гибрида, поскольку для этого используются и гомозиготные гены, т.к. при биологическом и механическом засорении гибрида может изменяться спектр любого изофермента.
Изоферментные маркеры в условиях отсутствия роботизированной техники для проведения ПЦР остаются востребованными в силу быстроты их определе- ния для паспортизации и для определения генетической чистоты коммерческих партий линий и гибридов подсолнечника. Но достичь с помощью только изофермент-ных маркеров 100 %-ной отличимости линий и гибридов не представляется возможным. Необходимо использование ПЦР-производных молекулярно-генетических маркеров и оптимизация их количества. В предыдущих работах было идентифицировано 17 инбредных линий коллекции ВНИИМК при помощи 10 SSR-локусов. Использование данных локусов позволило отличить генотипы друг от друга в 100 % случаев [19]. Использование трёх SSR и трёх SCAR-локусов для генотипирования коллекции 32 отечественных селекционных образцов подсолнечника позволило идентифицировать образцы лишь в 59 % случаев [13].
В результате анализа одиннадцати линий и четырех гибридов подсолнечника по восьми SSR-локусам были получены специфические и хорошо воспроизводимые фрагменты ДНК. Для каждого образца определены индивидуальные SSR-спектры, различающиеся числом ампликонов и их размерами на электрофореграммах (рис. 2).
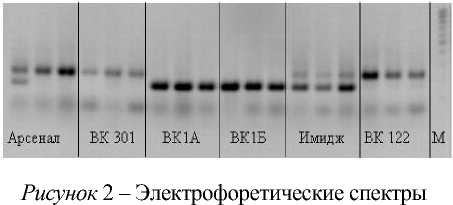
продуктов амплификации ДНК у линий и гибридов подсолнечника по локусу ORS 1796.
М – маркер молекулярного веса 100 пн
Количество аллелей для различных локусов варьировало от 1 до 2. Среднее число аллелей на локус составило 1,6 (с учетом мономорфных локусов) и 2 (без учета мономорфных локусов). Эти показатели совпадают с полученными ранее для других линий и гибридов подсолнечника [14; 19]. Три локуса не показали полиморфизма: IUB-6, ORS 553, RTS29. Остальные пять локусов показали по два аллельных состояния (табл. 3, 4).
Для обозначения аллельных вариантов использовали количество пар нуклеотидных оснований (пн) фрагмента ДНК. Гетерозиготные спектры, т.е. спектры, в которых присутствовали фракции двух разных аллелей, в таблицах обозначены двумя числами, разделенными косой чертой (/). При анализе электрофоретических спектров линий выявлена индивидуальность аллельного состава каждой из них, за исключением линий ВК 1 А ими и ВК 1 Б ими, которые, как было отмечено выше, являются стерильным и фертильным аналогами (табл. 3). Процент уникальности анализированной группы линий составил 91.
Таблица 3
Аллельные состояния микросателлитных локусов 11 линий подсолнечника
|
Генотип |
SSR-локус |
|||||||
|
ORS 509 |
ORS 595 |
IUB-6 |
ORS 1144 |
ORS 1796 |
ORS 553 |
RTS 29 |
ORS 1036 |
|
|
ВК 302 |
195* |
100 |
350 |
177 |
157 |
135 |
320 |
245 |
|
ВК 101 |
220 |
127 |
350 |
136 |
157 |
135 |
320 |
245 |
|
ВК 876 |
195 |
127 |
350 |
136 |
157 |
135 |
320 |
245 |
|
ВК 195 |
195 |
127 |
350 |
177 |
157 |
135 |
320 |
245 |
|
Р 96 |
220 |
127 |
350 |
177 |
157 |
135 |
320 |
245 |
|
ВК 930 |
220 |
127 |
350 |
177 |
232 |
135 |
320 |
255 |
|
RG 8 |
220 |
127 |
350 |
136 |
232 |
135 |
320 |
255 |
|
ВК 301 |
220 |
127 |
350 |
177 |
232 |
135 |
320 |
245 |
|
ВК 1 А ими |
220 |
127 |
350 |
136 |
157 |
135 |
320 |
245 |
|
ВК 1 Б ими |
220 |
127 |
350 |
136 |
157 |
135 |
320 |
245 |
|
ВК 22 |
195 |
127 |
350 |
177 |
232 |
135 |
330 |
255 |
|
* – количество пар нуклеотидных оснований (пн) аллеля SSR-локуса |
||||||||
Четыре исследованных гибрида имели отличающиеся по аллельному составу микросателлитных локусов генотипы (табл. 4).
Таблица 4
Аллельные состояния микросателлитных локусов четырех гибридов подсолнечника
|
Генотип |
SSR-локус |
|||||||
|
ORS 509 |
ORS 595 |
IUB-6 |
ORS 1144 |
ORS 1796 |
ORS 553 |
RTS 29 |
ORS 1036 |
|
|
Фактор |
195 ** / 220 |
100* |
350 |
136/ 177 |
157 |
135 |
320 |
245 |
|
Окси |
195 |
127 |
350 |
136/ 177 |
157 |
135 |
320 |
245 |
|
Арсенал |
220 |
127 |
350 |
136/ 177 |
232 |
135 |
320 |
245 |
|
Имидж |
195/ 220 |
127 |
350 |
136/ 177 |
157/2 32 |
135 |
320/ 330 |
245/25 5 |
|
* – количество пар нуклеотидных оснований (пн) аллеля гомозиготного SSR-локуса ** – количество пар нуклеотидных оснований (пн) аллелей SSR гетерозиготного локуса |
||||||||
Для гибрида Фактор характерно гетерозиготное состояние локусов ORS 509 и ORS 1144. Хотя родительские формы гибрида Фактор (линии ВК 101 и ВК 302) обладали отличающимися аллельными вариантами по локусу ORS 595, но в силу доминирования аллеля 100 пн у линии ВК 302 гибрид имеет по данному локусу лишь одно аллельное состояние – 100 пн. Гибриды Окси и Арсенал гетерозиготны по локусу ORS 1144. Гибрид Имидж обладал наибольшим количеством локусов в гетерозиготном состоянии: ORS 509, ORS 1144, ORS 1796, RTS 29 и ORS 1036. Для гибридов Имидж и Окси, у которых не было изоферментных локусов в гетерозиготном состоянии, выявлены SSR-локусы, по которым возможно определение уровня гибридности. Совокупный процент уникальности анализированных линий и гибридов составил 93.
Таким образом, с использованием пяти изоферментных и восьми SSR-локусов были составлены биохимические и молекулярно-генетические паспорта и установлена уникальность каждой инбредной линии (за исключением линий-аналогов) и гибридной комбинации. Процент уникальности линий, выявленный при помощи анализа изоферментных систем, составил 67, а при помощи микросател-литных локусов – 91. Совокупный процент уникальности линий и гибридов, выявленный при помощи анализа изо-ферментных систем, составил 75, при помощи микросателлитных локусов – 93. Установлено, что полиморфными и информативными для данной группы образцов являются четыре изоферментные системы: эстераза, малатдегидрогеназа, 6-фосфоглюконатдегидрогеназа, глюкозофосфатизомераза и пять микросателлит-ных локусов: ORS509, ORS 595, ORS1144, ORS1796, ORS 1036.
Список литературы Паспортизация новых линий и гибридов подсолнечника селекции ВНИИМК с помощью биохимических и молекулярных маркеров
- Вдовиченко Л.Д., Глазко В.И. Генетическая паспортизация сортов пшеницы с использованием ISSR-PCR маркеров//Сельскохозяйственная биология. -2007. -№ 3. -С. 33-37.
- Бобошина И.В., Боронникова С.В. Идентификация перспективных для Урала сортов пшеницы мягкой с использованием межмикросателлитного анализа полиморфизма ДНК//Фундаментальные исследования. -2013. -№ 6. -С. 92-97.
- Рамазанова С.А. Идентификация генотипов сои разного происхождения с использованием полиморфизма девяти микросателлитных локусов ДНК//Сб. статей 2-й междунар. конф. по сое: Современные проблемы селекции и технологии возделывания сои, Краснодар, 9-10 сентября. -2008.-С. 129-136.
- Супрун И.И., Токмаков С.В., Малюченко О.П., Ушакова Я.В., Бабаков А.В. Генотипирование сортов яблони российской селекции с использованием микросателлитных маркеров//Известия ТСХА. -2011. -Выл. 6.-С. 162-166.
- Гончарова Ю.К., Иванов А.Н., Князева КВ., Глазко В.И. Эстеразные спектры и адаптивная пластичность сортов риса//Доклады российской академии сельскохозяйственных наук. -2007. -№ 1. -С. 3-4.
- Cabezas J.A., Ibanez J„ Lijavetzky D., Velez D., Bravo G„ Rodriguez V. . A 48 SNP set for grapevine cultivar identification//Plant Biology. -2011. -№ 11.-P. 153.
- Саналатий A.B., Солоденко A.E., Сиволап Ю.М. Идентификация генотипов подсолнечника украинской селекции при помощи SSRP-анализа//Цитология и генетика. -2006. -Т. 40. -№ 4. -С. 31-37.
- Imerovski Dimitrijevic A., Miladinovic D„ Dedic D., Jocic.S'., Miklic V. Molecular profiles of sunflower lines resistant to broomrape (Orobanche cumana Wallr.)//International Symposium on Broomrape (Orobanche spp.) in Sunflower. Chisinau, Republic of Moldova. August 25-27. -2011. -P. 25.
- Усатое A.B., Маркин H.B., Горбаченко Ф.И., Федорова МА., Тихобаева В.Е., Горбаченко О.Ф., Азарин КВ. SSR-анализ геномной ДНК ЦМС-линий подсолнечника//Масличные культуры. Науч.-тех. бюл. ВНИИМК. -2011. -№ 1 (146-147). -С. 15-20.
- Moisan-Thiery М., Marhadour.S'., Kerlan М. С., Dessenne К, Perramant М., Gokelaere Т., Hingrat Y. Le. Potato cultivar identification using simple sequence repeats markers (SSR)//Potato Research. -2005. -Vol. 48. -Is 3^1. -P.191-200.
- Zhang L. S, Le Clerc V, Li S., Zhang D. Establishment of an effective set of simple sequence repeat markers for sunflower variety identification and diversity assessment//Canadian Journal of Botany. -2005. -83 (1). -P. 66-72.
- Гучетль C.3. Челюстникова T.A., Рамазанова С. А., Антонова Т.С. Молекулярногенетическая характеристика инбредных линий подсолнечника по изоферментным маркерам и ДНК профилям//Науч.-тех. бюл. ВНИИМК. -2004. -Вып. 2 (131). -С. 42-46.
- Гучетль С.З., Челюстникова Т.А., Арасла-нова Н.М., Антонова Т.С. SSR и SCAR генотипирование коллекции отечественных селекционных образцов подсолнечника, устойчивых и восприимчивых к расе Е Orobanche cumana Wallr.//Масличные культуры. Науч.-тех. бюл. ВНИИМК. -2012. -Вып. № 1 (150). -С. 20-26.
- Челюстникова ТА. Полиморфизм микро-сателлитных локусов в генотипах культурного и дикорастущего подсолнечника//Масличные культуры. Науч.-тех. бюл. ВНИИМК -2008. -Вып. 2(139). -С. 19-23.
- Туркав С.З., Лоскутов А.В., Губенко Т.П. Оценка генетической чистоты линий и гибридов подсолнечника с помощью изоферментных маркеров//Науч.-тех. бюл. ВНИИМК. -1996. -Вып. 117.-С. 33-37.
- Cardy B.S., Stuber C.W., Goodman М.М. Techniques for starch gel electrophoresis of enzymes for maize (Zea mays L.)//Institute of Statist. Mineograph. Series, 1317, North Carolina State Univers. -1981. -154 c.
- Vallejos C.E. Enzyme activity staining//Isozymes in plant genetics and breeding: Part A. -Amsterdam: Elsevier. -1983. -P. 469-516.
- Saghai-Maroof M.A., Soliman K.M., Jorgensen R.A., Allard R.W. Ribosomal DNA spacer-length polymorphisms in barley: mendelian inheritance, chromosomal location, and population dynamics//Proc. NAS USA. -1984. -Vol. 81. -P. 8014-8018.
- Antonova T.S., Guchetl S.Z., Tchelustnikova ТА., Ramazanova S.A. Development of marker system for identification and certification of sunflower lines and hybrids on the basis of SSR-analysis//Helia. -2006. -Vol. 29. -№ 45. -P. 63-72.
