Полиморфные варианты генов ферментов системы детоксикации ксенобиотиков CYP2Е1, GSTM1, GSTT1, EPHX1 как биомаркеры чувствительности к экспозиции побочными продуктами водоподготовки (на примере хлороформа)
")
Автор: Дроздова Елена Валентиновна, Колеснева Екатерина Владмировна, Сяхович Виталий Эдуардович, Долгина Наталья Алексеевна
Журнал: Анализ риска здоровью @journal-fcrisk
Рубрика: Медико-биологические аспекты оценки воздействия факторов риска
Статья в выпуске: 1 (41), 2023 года.
Бесплатный доступ
Установлено, что накопление хлороформа в организме и повышение его равновесных концентраций в крови экспонированного населения ассоциировано с наличием в генотипе полиморфизмов генов ферментов, участвующих в метаболизме побочных продуктов дезинфекции воды (A415G гена EPHX1, C1091T гена CYP2E1, нулевых мутаций генов глутатионтрансферазы GSTT1 и GSTM1) (р
Гены cyp2е1, gstm1, gstt1, ephx1, продукты дезинфекции воды, питьевая вода, полиморфизм генов, биомониторинг, оценка риска здоровью, биомаркеры чувствительности
Короткий адрес: https://sciup.org/142237421
IDR: 142237421 | УДК: 614.777-047.36 | DOI: 10.21668/health.risk/2023.1.15
Polymorphisms of xenobiotic metabolism enzyme genes CYP2E1, GSTM1, GSTT1, EPHX1 as biomarkers of sensitivity to exposure to water disinfection byproducts (using chloroform as an example)
Chloroform accumulation in the body and the increase in its steady-state concentrations in blood of exposed people have been established to be associated with polymorphisms of enzyme genes in a genotype involved in metabolism of water disinfection byproducts (A415G of EPHX1 gene, C1091T of CYP2E1 gene, zero mutations of GSTT1 and GSTM1 genes) (p
Текст научной статьи Полиморфные варианты генов ферментов системы детоксикации ксенобиотиков CYP2Е1, GSTM1, GSTT1, EPHX1 как биомаркеры чувствительности к экспозиции побочными продуктами водоподготовки (на примере хлороформа)
в относительно низких и следовых концентрациях, их одновременное воздействие определяет риски здоровью населения при водопользовании, особенно при длительном поступлении [1–11]. Использование воды, содержащей смесь тригалометанов, повышает риски развития нарушений течения беременности у женщин (задержка внутриутробного развития плода, снижение массы тела новорожденных, преждевременные роды, врожденные дефекты развития плода) [1, 6]. По степени доказанности канцерогенного действия на человека хлороформ, бромоформ и ди-хлорбромметан Международным агентством по изучению рака (МАИР) отнесены к группе 2B (возможный канцероген для человека) на основании доказательств в экспериментах на животных [1, 2, 7–9]. Эпидемиологические исследования показали связь между воздействием тригалометанов и раком мочевого пузыря, раком прямой кишки [1, 10, 11]. Учитывая новые данные об опасности тригаломета-нов, на международном уровне наметилась тенденция к ужесточению нормативов для содержания данной группы ППД в питьевой воде. Однако пересмотр национальных нормативов требует формирования надежной доказательной базы с учетом условий водопользования в республике и особенностей чувствительности популяции к экзогенным загрязнителям, что может быть достигнуто за счет применения ряда методических подходов [12].
Из-за сложного химического состава смесей ППД и множественных путей поступления оценка фактического воздействия тригалометанов на организм человека является очень сложной задачей. Повышение точности и объективности количественной оценки рисков здоровью населения идет в том числе по пути применения данных о внутренней экспозиции – содержании загрязнителей (их метаболитов) во внутренних средах организма вещества [13–16]. Доказано, что уровни содержания хлороформа в крови могут расцениваться как биомаркеры экспозиции тригалометанами с водой. Концентрация хлороформа в крови повышается сразу после сеанса душа / ванной, мытья посуды вручную, употребления приготовленных на водопроводной воде горячих напитков (наибольшие уровни отмечаются после душа / ванной) и быстро снижается [17–20]. Однако и через 8 ч в крови можно обнаружить незначительные количества хлороформа. Постепенное его высвобождение из депо в жировой ткани при интенсивном воздействии (частый прием душа или ванной и т.д.) приводит к формированию «установившихся» концентраций хлороформа в крови (steady-state level ). Исследование образца крови без привязки к факту водопользования позволяет установить эти концентрации и более точно оценить внутреннюю экспозицию для последующей оценки рисков здоровью [21].
В то же время, помимо интенсивности воздействия и массы тела, на уровни содержания тригало-метанов в крови человека влияет комплекс факто- ров, важнейшим из которых является активность системы детоксикации ксенобиотиков. Межиндивидуальные различия в способности к биотрансформации ксенобиотиков (их метаболитов) могут быть обусловлены полиморфизмом генов, кодирующих как сами ферменты, так и рецепторы и / или факторы транскрипции, регулирующие их экспрессию. Кроме того, полиморфизмы в нескольких областях генов могут привести к изменению сродства к лиганду, активности трансактивации или уровня экспрессии рецептора, что впоследствии влияет на экспрессию генов-мишеней [22–24]. Как следствие, замедление выведения токсичных веществ и / или увеличение образования токсичных метаболитов, их накопление в организме повышают вероятность реализации негативных биологических эффектов и, соответственно, развития экологически детерминированных хронических неинфекционных заболеваний. Таким образом, наличие в генотипе полиморфизмов генов системы детоксикации ксенобиотиков может быть основой различий в индивидуальной восприимчивости к воздействию химических факторов окружающей среды, а аномальные аллели генов (полиморфизмы) ферментов, принимающих участие в метаболизме ксенобиотиков, являясь предикторами риска развития указанных выше заболеваний, могут использоваться как маркеры чувствительности организма к воздействию токсикантов [24–27].
Цитохром Р450 2E1 (CYP2E1) – основной фермент, катализирующий процесс детоксикации тригалометанов в организме людей и крыс. Относительная значимость второстепенных путей метаболизма продуктов деструкции – фосгена – зависит от наличия глутатиона, других тиолов и других нуклеофильных соединений (гистидин и цистеин). Восстановленный глутатион способен элиминировать практически все метаболиты хлороформа, образующиеся при не слишком высоких концентрациях хлороформа. Баланс между окислительным и восстановительным путями детоксикации тригалометанов (и хлороформа в частности) зависит от вещества, ткани, дозы и выраженности окислительного стресса. Для гена CYP2E1 наиболее часто рассматриваются тесно сцепленные полиморфизмы по рестрикционным эндонуклеазам PstI/RsaI, при которых мутантный аллель способствует повышенной транскрипционной и ферментативной активности. Частота встречаемости этого аллеля отличается в разных популяциях: у азиатов 6 % жителей являются носителями гомозиготного аллеля, 35 % – гетерозиготны, тогда как среди европейцев гетерозиготами является менее 6 % индивидуумов.
Полиморфизм локализованного в хромосоме 1 гена GSTM1 (глутатион-S-трансфераза класса µ) обусловлен наличием двух аллелей: функционально активного GSTM(+) и неактивного «нулевого» GSTM(-), имеющего обширную делецию, из-за чего синтез белкового продукта не происходит вообще.
Локализованный на хромосоме 22 ген GSTT1 (глу-татион-S-трансфераза (GST) тета-1) также представлен двумя аллелями: функционально активным GSTT1(+) и неактивным «нулевым» GSTТ1(-), соответствующим частичной или полной делеции, приводящей к снижению активности белка или к его отсутствию. Зарубежными учеными L.C. Backer et al. (2008) было установлено, что участники исследования с GSTT1(-) имели более высокий уровень хлороформа в крови после душа, чем GSTT1(+)-участники [20]. Исследование Kenneth P. Cantor et al. (2010) подтвердило гипотезу о связи рака мочевого пузыря у человека с генетическим полиморфизмом GSTM1, GSTZ1 [28]. В ряде статей приводятся доказательства задержки роста плода при наличии у матерей генетических полиморфизмов генов: CYP2E1, MTHFR [29], CYP2E1 и GSTZ1 [30], CYP2E1 [31]. В кросс-секционном исследовании P. Yang et al. (2016) показана связь между качеством спермы у мужского населения одной из территорий в условиях экспозиции ТГМ с водой и наличием полиморфизмов генов CYP2E1 и GSTZ1 и GSTT1 [32]. Распространенность нулевых генотипов GSTM1(-) и GSTT1(-) среди лиц европеоидной расы – 40–50 и 10–20 % соответственно [33].
Для гена микросомальной эпоксидгидролазы, являющейся важным компонентом процессов метаболизма чужеродных соединений (EPHX1), известны два функционально значимых полиморфизма, приводящие к изменению свойств фермента: в 3-м экзоне (Т337С – замена тирозина на гистидин в 113-м положении (Tyr113His)) и в 4-м экзоне (A415G – замена гистидина на аргинин в 139-м положении (His139Arg)). Полиморфизм Т337С отвечает за снижение активности фермента на 50 % («медленный» аллель), а полиморфизм A415G – за повышение активности примерно на 25 % («быстрый» аллель). Ускоренное превращение производных эпоксидов в высокоактивные метаболиты приводит к повреждению ДНК, что способствует увеличению количества хромосомных аберраций и является одной из причин развития ряда патологий.
В Республике Беларусь исследования уровней ТГМ в крови населения на территориях с водоснабжением из поверхностных источников, а также влияния генетической вариабельности популяции на их формирование до настоящего времени не проводились. Рабочая гипотеза нашего исследования состояла в том, что наличие в генотипе полиморфизмов генов системы детоксикации ксенобиотиков, принимающих участие в метаболизме хлороформа, может иметь следствием формирование потенциально более высоких уровней его содержания в биологических средах экспонированного населения при длительном воздействии, что обусловливает в конечном счете более высокую вероятность реализации рисков, ассоциированных с неблагоприятным воздействием побочных продуктов дезинфекции на организм.
Цель исследования – изучить влияние полиморфизма генов ферментов CYP2E1, GSTM1, GSTT1, EPHX1 на метаболизм побочных продуктов дезинфекции и обосновать биомаркеры индивидуальной чувствительности экспонированного населения к воздействию тригалометанов (на примере хлороформа).
Материалы и методы. В рамках работы были сформированы две группы добровольцев из числа населения г. Минска: группа наблюдения – экспонированное население, проживающее в Московском и Фрунзенском районах, снабжаемых водой из поверхностного источника, прошедшей дезинфекцию хлорированием (150 человек); контрольная группа (группа сравнения) – неэкспонированное население, проживающее в Первомайском районе, снабжаемом водой из подземных источников без применения хлорирования (47 человек). Группы были сопоставимы по возрасту (18–40 лет) и полу.
Содержание хлороформа в водопроводной воде на территории наблюдения в период, предшествующий исследованию, и во время исследования было в пределах нормативных требований – от 0,49 до 0,52 ПДК (ПДК ≤ 0,2 мг/дм3). На территории сравнения хлороформ в воде разводящей сети в пределах чувствительности метода не определялся (< 0,0125 мг/дм3). Определение хлороформа в воде проводили методом газовой хроматографии ( LOD для хлороформа 0,0125 мг/дм3).
Проведен забор периферической крови у добровольцев на базе поликлинических медицинских учреждений г. Минска в весенний период в утренние часы. Все стадии исследования соответствовали этическим нормам, у всех участников исследования было получено информированное согласие на участие в исследованиях, а также проведено анкетирование по индивидуальному водопользованию.
В качестве биомаркеров экспозиции во всех образцах крови определяли концентрации хлороформа. Исследование проводилось на базе учреждения здравоохранения «Национальная антидопинговая лаборатория» методом газовой хроматографии-масс-спектрометрии низкого разрешения, разработанным в рамках настоящего задания (СОП LM 174-2020 «Метод определения тригалометанов (хлороформа) в биологических средах»), на газовом хроматографе AGILENT 7890 с масс-спектрометрическим детектором типа «тройной квадруполь» AGILENT 7000 (Thermo Fisher Scientific, США) [34].
Также генотипы всех добровольцев были исследованы на наличие полиморфизмов A415G гена EPHX1, C1091T гена CYP2E1 и нулевых мутаций генов глутатион S-трансферазы класса µ и θ (GSTT1(-) и GSTM1(-)). Выбор генов-кандидатов для исследования основывался на особенностях метаболизма тригалометанов (хлороформа) в организме. Генотипирование проводилось на базе Республиканского унитарного предприятия «Научно-практический центр гигиены». ДНК выделяли из образцов крови с помощью коммерческого набора для выделения ДНК «Нуклеосорб», комплектация «В» производства ОДО «Праймтех» (Беларусь) согласно протоколу фирмы-изготовителя. Для установления генотипов по целевым локусам применяли метод полимеразной цепной реакции (ПЦР) в реальном времени с использованием прибора «С1000» (BioRad, США). Полученные результаты в последующем анализировались с применением соответствующего программного обеспечения. Информация о характеристиках полиморфизмов, локализации анализируемых генов и используемых ферментах рестрикции приведена в табл. 1.
Статистический анализ. Различия в распределении частот генотипов и аллелей среди лиц из группы наблюдения и лиц в группе сравнения анализировались с помощью критерия χ2. Характер распределения наблюдаемых частот генотипов и аллелей генов в популяции оценивали на соответствие уравнению Харди – Вайнберга. Качественные показатели представлены в виде абсолютного значения и доли. При описании количественных признаков (содержание хлороформа в крови) использовались общепринятые показатели: max – максимальные значения, min – минимальные значения; Ме – медиана, значения верхнего и нижнего квартилей (интерквартильный размах) – [Р25; Р75], 10, 90-го и 95-го процентилей (Р10, Р90, Р95), доверительный интервал – 95 % ДИ. При анализе количественных данных использовались методы статистической обработки с учетом вида распределения данных: при сравнении несвязанных выборок по изучаемому признаку применялся t -тест для независимых выборок, U- критерий Манна - Уитни. При сравнении показателя в нескольких независимых группах применяли непараметрический критерий Краскела – Уоллиса ( Н ). Для каждого полиморфизма и их комбинаций были рассчитаны отношения шансов (ШО, RO) с учетом 95%-ного доверительного интервала выявления в крови более высоких (≥ Р75) концентраций хлороформа у лиц с определенными полиморфизмами в сравнении с лицами без полиморфизмов.
В исследовании значение р < 0,05 считали статистически значимым. Анализ данных проводился с использованием стандартного пакета прикладных статистических программ Statistica 12.0 и Microsoft Office Excel.
Распределение генотипов и аллелей полиморфизмов исследованных генов среди добровольцев в двух группах представлено в табл. 2, оно соответствовало равновесию Харди – Вайнберга.
Полученные результаты изучения распределения полиморфных вариантов генов биотрансформации ксенобиотиков в группах наблюдения и сравнения в целом согласуются с литературными данными о распределении полиморфных вариантов генов системы метаболизма ксенобиотиков в европейской популяции. При изучении распределения частоты генотипов полиморфного локуса C1091T гена CYP2E1 гетерозиготные генотипы СТ были выявлены у 10 % лиц из группы наблюдения и у 4,3 % лиц из группы сравнения, остальные генотипы индивидуумов по данному локусу соответствовали гомозиготному генотипу дикого типа СС. Распределение нулевых генотипов GSTM(-) и GSTT1(-) составило соответственно 36,7 и 23,3 % в группе наблюдения и 36,2 и 17,0 % в группе сравнения. Анализ распределения полиморфизма 4-го экзона А415G гена EPHX1 показал, что частота встречаемости полиморфного аллеля G в опытной группе составляет 15,7 %, мутантных гомозигот GG – 9,3 %, гетерозиготного генотипа AG – 12,7 %, в то время как в контрольной группе – 8,7; 4,2 и 12,8 % соответственно.
58,7 % лиц из группы наблюдения и 51,1 % из группы сравнения имели мутантный гомо- и гетерозиготный генотипы хотя бы по одному из четырех генов: GSTM1, GSTT1, EPHX1, CYP2E1. В группе сравнения полиморфизм одного из четырех генов отмечен в 32,9 % случаев, полиморфизмы двух генов – в 18,7 %, трех генов – в 7,3 %. Встречаемость одновременно полиморфизмов нескольких генов в группах наблюдения и сравнения составила соответственно для GSTM1+EPHX1 – 6,7 и 8,5 %, GSTM1+GSTT1 – 5,3 и 4,3 %, GSTT1+EPHX1 – 2,7 и 0,0 %, для GSTT1+CYP2E1 – 2,0 и 2,1 %, EPHX1+CYP2E2 – 2,7 и 2,1 %, GSTM1+GSTT1+EPHX1 – 2,7 и 2,1 %, GSTM1+GSTT1+CYP2E1 – 2,0 и 2,1 %, GSTT1+ EPHX1+CYP2E1 – 2,7 и 0,0 %.
Таблица 1
Последовательности праймеров и ферменты рестрикции, используемые при ПЦР-анализе полиморфизма целевых генов-кандидатов
|
Ген, локализация |
Полиморфизм |
Последовательность праймеров, (5’ › 3’) |
Длина продукта, пн |
Фермент рестрикции |
Аллель, пн |
|
CYP2Е1 10q24.3 |
5`-регуляторная область С1091T |
F CCAGTCGAGTCTACATTGTCA R TTCATTCTGTCTTCTAACTGG |
411 |
RsaI |
C (412) T (351, 61) |
|
GSTM1 1q13.3 |
Делеция |
F GAACTCCCTGAAAAGCTAAAGC R GTTGGGCTCAAATATACGGTGG |
219 |
- |
норма (219) делеция (0) |
|
GSTT1 22q11.2 |
Делеция |
F TCACCGGATCATGGCCAGCA R TTCCTTACTGGTCCTCACATCTC |
459 |
- |
норма (459) делеция (0) |
|
EPHX1 1g42.1 |
4-й экзон А415G (His139Arg) |
F ACATCCACTTCATCCACGT R ATGCCTCTGAGAAGCCAT |
210 |
RsaI |
A (210) G (163, 47) |
Таблица 2
Распределение аллелей и генотипов полиморфизмов A415G гена EPHX1 (His139Arg), C1091T гена CYP2E1, GSTT1(-) и GSTM1(-) в группах наблюдения и сравнения
|
Генотипы/ аллели* |
Частота генотипов, % |
Значение р (для различных групп) |
|
|
Группа наблюдения (опытная) |
Группа сравнения (контроль) |
||
|
C1091T CYP2E1 |
|||
|
CC |
90,0 |
95,7 |
0,225 |
|
CT |
10,0 |
4,3 |
|
|
ТТ |
0,0 |
0,0 |
|
|
C |
95,0 |
97,9 |
0,393 |
|
T |
5,0 |
2,1 |
|
|
GSTT1 |
|||
|
n |
76,7 |
83,0 |
0,361 |
|
del |
23,3 |
17,0 |
|
|
GSTM1 |
|||
|
n |
63,3 |
63,8 |
0,951 |
|
del |
36,7 |
36,2 |
|
|
A415G EPHX1 |
|||
|
AA |
78,0 |
83,0 |
0,461 |
|
GA |
12,7 |
12,8 |
0,986 |
|
GG |
9,3 |
4,2 |
0,263 |
|
A |
84,3 |
91,3 |
0,228 |
|
G |
15,7 |
8,7 |
|
П р и м е ч а н и е : * AA, СС – гомозиготный генотип дикого типа; AG, CT – гетерозиготный генотип; GG, ТТ – гомозиготный мутантный генотип; A, С – аллель дикого типа; G, Т – мутантный аллель; n – дикий тип гена; del – гомозиготная делеция.
Статистически значимые различия встречаемости относительных частот мутантного гомозиготного, гетерозиготного и нормального гомозиготного генотипа по изученным генам в группах наблюдения и контроле отсутствовали по всем парам сравнения ( р > 0,05), что свидетельствует о сопоставимости сформированных выборок по группам.
Результаты и их обсуждение. В 100 % проб крови обеих групп был выявлен хлороформ: в группе наблюдения – в концентрациях от 0,03 до 0,54 нг/мл, Р95 – 0,27 нг/мл, в контрольной группе – от 0,004 до 0,37 нг/мл, Р95 – 0,13 нг/мл. Различия содержания в крови хлороформа у экспонируемого и неэкспонируемого населения (через воду) статистически значимы ( U = 2336, р < 0,01) [34]. Выявленные концентрации хлороформа в крови не превышали (за исключением одного образца) реперные концентрации (0,0004 мг/л), предложенные российскими учеными по повышению содержания в крови аспартатаминотрансферазы [16].
На следующем этапе оценивали наличие ассоциаций между наличием полиморфизмов соответствующих генов, кодирующих ферменты системы детоксикации ксенобиотиков, и уровнями хлороформа в крови населения. Далее в статье описаны выявленные ассоциации и закономерности только по группе наблюдения, поскольку статистически значимые различия между распределением аллелей в группах наблюдения и сравнения отсутствовали, а установленные статистически значимые отличия по основным ассоциациям полиморфизмов изучаемых генов с концентрациями хлороформа в крови лиц контрольной группы соответствовали тенденциям в группе наблюдения.
Для анализа ассоциаций результаты определения концентраций хлороформа в крови добровольцев группы наблюдения были обобщены по 42 условным подгруппам (вариантам) в зависимости от наличия / отсутствия полиморфизмов изучаемых генов: отсутствие в генотипе изучаемых полиморфизмов генов (подгруппа № 2), наличие в генотипе изучаемых полиморфизмов (№ 3), наличие в генотипе одного из изучаемых полиморфизмов генов (моно) (№ 4–7), наличие в генотипе полиморфизма гена в сочетании с полиморфизмами других генов (№ 8–11), наличие в генотипе полиморфизмов двух или трех генов в различных сочетаниях (№ 12–22), отсутствие в генотипе полиморфизмов конкретных генов (№ 23–26) или их определенных сочетаний (№ 27–42). Учитывая, что лишь в части подгрупп распределение концентраций соответствовало нормальному, в дальнейшем для анализа данных применялись непараметрические методы. По каждой подгруппе были рассчитаны число проб и их доля от общего числа проб в группе наблюдения, концентрации – max, min, Ме , Р25, Р75, Р10, Р90, а также число проб в подгруппе (абсолютные значения и доля в % от проб в подгруппе) с содержанием хлороформа ≥ значений Р75 и Р90, ≤ значений Р10, Р25, Р75 и Р90 для группы наблюдения в целом.
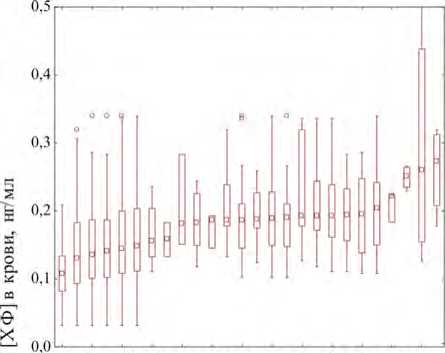
Обобщенные данные по подгруппам по содержанию хлороформа в крови добровольцев группы наблюдения в зависимости от наличия (отсутствия) полиморфизмов представлены в табл. 3 (26 подгрупп, представляющих наибольший исследовательский интерес), а визуализация результатов в виде диаграммы размаха по основным 19 подгруппам – на рис. 1.
Таблица 3
Уровни хлороформа в крови волонтеров в группе наблюдения, нг/мл
|
№ п/п |
Подгруппа по наличию/отсутствию полиморфизмов (пояснение) |
N набл. |
% от всех проб |
Me [Р25 - Р75] |
Р10 |
Р90 |
Число проб (%) с концентрацией хлороформа более (менее) соответствующего процентиля по группе наблюдения в целом |
|||
|
≥ Р75 |
≥ Р90 |
< Р10 |
< Р25 |
|||||||
|
1 |
В целом группа наблюдения |
150 |
100,0 |
0,1488 [0,1118 ÷ 0,2035] |
0,0819 |
0,2392 |
25,3 |
10,0 |
10,0 |
25,3 |
|
2 |
Нет полиморфизмов |
62 |
41,3 |
0,1082 [0,0822 ÷ 0,1341] |
0,0628 |
0,1568 |
4,8 |
0,0 |
24,2 |
56,5 |
|
3 |
Есть полиморфизмы |
88 |
58,7 |
0,1892 [0,1495 ÷ 0,2281] |
0,1310 |
0,2660 |
60,2 |
17,0 |
0,0 |
23,9 |
|
4 |
Полиморфизм GSTM1 (моно) |
30 |
20,0 |
0,1903 [0,1480 ÷ 0,2105] |
0,1279 |
0,2351 |
30,0 |
6,7 |
0,0 |
26,7 |
|
5 |
Полиморфизм GSTT1 (моно) |
9 |
6,0 |
0,1826 [0,1497 ÷ 0,2257] |
0,1178 |
0,2440 |
44,4 |
11,1 |
0,0 |
22,2 |
|
6 |
Полиморфизм EPHX1 (моно) |
8 |
5,3 |
0,1951 [0,1387 ÷ 0,2478] |
0,1081 |
0,2862 |
50,0 |
37,5 |
0,0 |
37,5 |
|
7 |
Полиморфизм CYP2E1 (моно) |
2 |
1,3 |
0,1581 [0,1333 ÷ 0,1829] |
0,1333 |
0,1829 |
0,0 |
0,0 |
0,0 |
50,0 |
|
8 |
Полиморфизм GSTM1 (не только моно) |
55 |
36,7 |
0,1869 [0,1455 ÷ 0,2105] |
0,1272 |
0,2387 |
30,9 |
9,1 |
0,0 |
27,3 |
|
9 |
Полиморфизм GSTT1 (не только моно) |
35 |
23,3 |
0,1930 [0,1711 ÷ 0,2440] |
0,1451 |
0,3061 |
48,6 |
28,6 |
0,0 |
14,3 |
|
10 |
Полиморфизм EPHX1 (не только моно) |
33 |
22,0 |
0,2040 [0,1494 ÷ 0,2422] |
0,1310 |
0,3061 |
54,5 |
30,3 |
0,0 |
24,2 |
|
11 |
Полиморфизм CYP2E1 (не только моно) |
15 |
10,0 |
0,1869 [0,1778 ÷ 0,2385] |
0,1455 |
0,3061 |
40,0 |
20,0 |
0,0 |
13,3 |
|
12 |
Полиморфизмы GSTM1+GSTT1 |
8 |
5,3 |
0,1873 [0,1745 ÷ 0,2268] |
0,1243 |
0,2586 |
37,5 |
12,5 |
0,0 |
12,5 |
|
13 |
Полиморфизмы GSTM1+EPHX1 |
10 |
6,7 |
0,1556 [0,1326 ÷ 0,2039] |
0,1219 |
0,2230 |
30,0 |
0,0 |
0,0 |
40,0 |
|
14 |
Полиморфизмы GSTT1+EPHX1 |
4 |
2,7 |
0,2512 [0,2346 ÷ 0,2643] |
0,2294 |
0,2660 |
100,0 |
75,0 |
0,0 |
0,0 |
|
15 |
Полиморфизмы GSTT1+CYP2E1 |
3 |
2,0 |
0,1819 [0,1514 ÷ 0,2832] |
0,1514 |
0,2832 |
33,3 |
33,3 |
0,0 |
0,0 |
|
16 |
Полиморфизмы EPHX1+CYP2E1 |
3 |
2,0 |
0,2222 [0,1839 ÷ 0,2237] |
0,1839 |
0,2237 |
66,7 |
0,0 |
0,0 |
0,0 |
|
17 |
Полиморфизмы GSTM1+GSTT1+EPHX1 |
4 |
2,7 |
0,2597 [0,1550 ÷ 0,4383] |
0,1272 |
0,5401 |
50,0 |
50,0 |
0,0 |
25,0 |
|
18 |
Полиморфизмы GSTM1+GSTT1+CYP2E1 |
3 |
2,0 |
0,1869 [0,1455 ÷ 0,1930] |
0,1455 |
0,1930 |
0,0 |
0,0 |
0,0 |
33,3 |
|
19 |
Полиморфизмы GSTT1+EPHX1+CYP2E1 |
4 |
2,7 |
0,2723 [0,2082 ÷ 0,3128] |
0,1778 |
0,3195 |
75,0 |
75,0 |
0,0 |
0,0 |
|
20 |
Полиморфизмы более 1 гена |
39 |
26,0 |
0,1930 [0,1617 ÷ 0,2385] |
0,1325 |
0,3061 |
46,2 |
23,1 |
0,0 |
17,9 |
|
21 |
Полиморфизмы 2 генов |
28 |
18,7 |
0,1944 [0,1565 ÷ 0,2325] |
0,1325 |
0,2626 |
46,4 |
17,9 |
0,0 |
17,9 |
|
22 |
Полиморфизм 3 генов |
11 |
7,3 |
0,1930 [0,1778 ÷ 0,3195] |
0,1455 |
0,3366 |
45,5 |
36,4 |
0,0 |
18,2 |
|
23 |
Нет полиморфизмов GSTM1 |
95 |
63,3 |
0,1310 [0,0935 ÷ 0,1829] |
0,0718 |
0,2398 |
22,1 |
10,5 |
15,8 |
63,2 |
|
24 |
Нет полиморфизма GSTT1 |
115 |
76,7 |
0,1363 [0,1013 ÷ 0,1872] |
0,0773 |
0,2222 |
18,3 |
4,3 |
13,0 |
57,4 |
|
25 |
Нет полиморфизма EPHX1 |
117 |
78,0 |
0,1414 [0,1025 ÷ 0,1869] |
0,0773 |
0,2225 |
17,1 |
4,3 |
12,8 |
57,3 |
|
26 |
Нет полиморфизма CYP2E1 |
135 |
90,0 |
0,1446 [0,1084 ÷ 0,2000] |
0,0812 |
0,2378 |
23,7 |
8,9 |
11,1 |
50,4 |
У экспонированных лиц из группы наблюдения, в генотипе которых присутствовал полиморфизм хотя бы одного изучаемого гена, хлороформ определялся в концентрациях от 0,1025 до 0,5401 нг/мл, Ме = 0,1892 нг/мл [0,1495; 0,2281], Р90 – 0,2660 нг/мл. У экспонированных лиц без изучаемых полиморфизмов генов в генотипе концентрации хлороформа находились в диапазоне от 0,0321 до 0,2087 нг/мл, Ме = 0,1082 нг/мл [0,0822; 0,1341], Р90 – 0,1568 нг/мл. Оценка достоверности установленных различий в содержании хлороформа в крови между подгруппами с наличием полиморфизмов генов и их комбинаций (№ 3–22) и подгруппой с отсутствием полиморфизмов (подгруппа № 2) по U-критерию Манна - Уитни показала, что различия статистически значимы (р < 0,01) для всех групп, за исключением подгруппы № 7 «полиморфизм CYP2E1 (моно)» (двустороннее р > 0,05), что может быть обусловлено малочисленностью данной группы. Дополнительно проведена оценка значимости различий между подгруппами с наличием полиморфизмов (№ 3–22) и подгруппами с отсутствием соответствующих полиморфизмов или их комбинаций (подгруппы № 23–26 и еще 16 подгрупп с отсутствием комбинаций полиморфизмов), различия статистически значимы для подгрупп № 3–6, 8–11, 14, 19–22 (р < 0,05) и статистически не значимы для подгрупп № 7, 12–13, 15–18 (р > 0,05).
Сравнение различий между 12 основными подгруппами (№ 2, 4–7, 12–19) методом Краскела – Уоллиса и медианным тестом показало их статистическую значимость: Н (12, N+150) = 80,5, р < 0,01, χ2 = 63,5, сс = 12, р < 0,01 (с поправкой Бонферрони). Попарные сравнения 16 основных подгрупп (№ 4–19) с использованием теста Манна – Уитни (всего 99 пар сравнения) позволили установить статистически
2 23 24 25 26 I 13 7 15 5 18 II 8 12 3 4 22 9 20 21 6 1016 1417 19
Подгруппа
° Медиана Q 25%-75% I Размах без выбр, о Выбросы • Крайние точки
Рис. 1. Диаграмма размаха для содержания хлороформа в крови волонтеров группы наблюдения в зависимости от наличия изучаемых полиморфизмов генов ферментов системы детоксикации ксенобиотиков CYP2E1, GSTM1, GSTT1, EPHX1 (подгруппы приведены в последовательности, отражающей возрастание значений медиан концентраций)
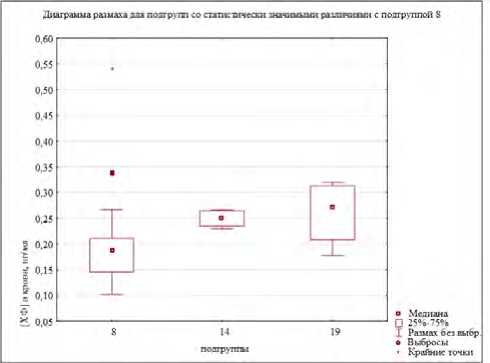
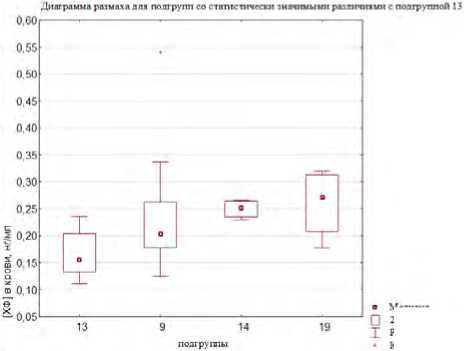
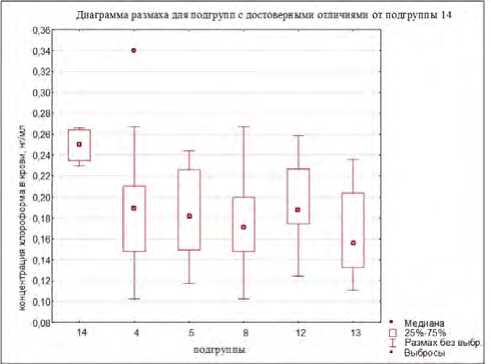
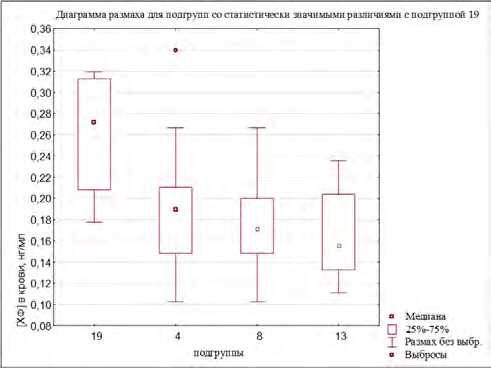
значимые различия между уровнями содержания хлороформа в крови волонтеров в следующих подгруппах: № 4 и 14 ( U = 10; Z = – 2,66; p < 0,01), № 4 и 19 ( U = 22; Z = – 2,00; p < 0,05), № 5 и 14 ( U = 3; Z = – 2,24; p < 0,05), № 12 и 14 ( U = 3; Z = – 2,12; p < 0,05), № 13 и 14 ( U = 1; Z = – 2,62; p < 0,01), № 13 и № 19 ( U = 4; Z = – 2,19; p < 0,05), № 13 и 9 ( U = 100; Z = – 2,03; p < 0,05), № 8 и 14 ( U = 22; Z = – 2,64; p < 0,01), № 8 и 19 ( U = 44; Z = – 1,97; p < 0,05). При этом уровни содержания хлороформа в крови (по медиане) были ниже в подгруппе № 4 (полиморфизм GSTM1 (моно)) на 32 и 43,1 % в сравнении с подгруппами № 14 (полиморфизмы GSTT1+EPHX1) и № 19 (полиморфизмы GSTT1+ EPHX1+CYP2E1) соответственно, ниже в группе – № 13 (полиморфизмы GSTM1+EPHX1) – на 24,1; 61,5 и 75,0 %, чем в подгруппах № 9 (полиморфизм GSTT1 (не только моно)), № 14 (полиморфизмы GSTT1+EPHX1) и № 19 (полиморфизмы GSTT1+ EPHX1+CYP2E1) соответственно, а также в подгруппе № 8 (полиморфизм GSTM1 (не только моно)) – на 34,4 и 45,7 %, чем в подгруппах № 14 и 19. Концентрации хлороформа в крови экспонированных лиц выше в подгруппе № 14 в сравнении с подгруппами № 4, 5, 12, 13 и 8 от 23,2 до 38,1 %, а также в подгруппе № 19 в сравнении с подгруппами № 4, 8 13 – на 30,2–42,9 % (рис. 2).
Проведен анализ различий между уровнями содержания хлороформа в крови в зависимости от наличия / отсутствия полиморфизма конкретного гена (результаты представлены на рис. 3). Наличие полиморфизмов генов GSTT1 и EPHX1, а также их одновременное присутствие отражаются на уровнях концентраций в сторону их увеличения (подгруппы № 10, 14, 17, 19).
Выполнен сравнительный анализ распределения проб с различными уровнями содержания хлороформа в крови (< P10, < P25, ≥ P75, ≥ P90 группы наблюдения) в подгруппах. Показано, что к подгруппе № 2 «нет полиморфизмов» относились все пробы с содержанием хлороформа в крови < Р10 (0,0819 нг/мл) (24,2 % проб в подгруппе) и 92,1 % проб с содержанием хлороформа < P25 (0,1118 нг/мл) (56,5 % по подгруппе). В то время как в подгруппе № 3 «есть полиморфизмы» доля проб с содержанием хлороформа < P25 составила в 2 раза меньше – 23,9 %.
В группе наблюдения из проб крови с содержанием хлороформа ≥ верхнего квартиля (Р75) (0,2035 нг/мл) 44,7 % приходилось на лиц с полиморфизмами генов GSTT1 или GSTM1, 47,4 % – с полиморфизмами EPHX1, 15,8 % – CYP2E1, и только в 7,9 % изучаемые полиморфизмы отсутствовали. Наибольший удельный вес проб с содержанием хлороформа ≥ Р75 отмечался в подгруппах № 5 «полиморфизм GSTT1 (моно)» (44,4 %), № 9 «полиморфизм GSTT1 (не только моно)» – 48,6 %, № 6 «полиморфизм EPHX1 (моно))» и № 17 «полиморфизмы GSTM1+GSTT1+EPHX1» – 50 %, № 10 «полиморфизм EPHX1 (не только моно)» – 54,5 %, № 3 «есть полиморфизмы» – 60,2 %, № 16 «полиморфизмы EPHX1+CYP2E1» – 66,7 %, № 19 «GSTT1+EPHX1+ CYP2E1» – 75 %, № 14 «полиморфизмы GSTT1+ EPHX1» – 100 %. При этом во всех подгруппах с долей проб ≥ Р75 50 % и более в генотипах присутствовали полиморфизмы гена EPHX1.
Все пробы с содержанием хлороформа в крови ≥ Р90 (0,2392 нг/мл) соответствовали лицам с наличием в генотипе полиморфизмов, из них 66,7 % имели полиморфизмы генов GSTM1 или EPHX1, 33,3 % – полиморфизмы GSTT1, 20,0 % – CYP2E1 (соответственно 75 % проб подгрупп № 13 «GSTT1+EPHX1» и № 19 «GSTT1+EPHX1+CYP2E1», 50 % проб подгруппы № 17 «GSTM1+GSTT1+EPHX1», 37,5 % проб подгруппы № 6 «полиморфизм EPHX1 (моно)»).
Расчет отношений шансов ( OR ) для наблюдаемой популяции показал (табл. 4), что для экспонированных лиц с наличием в генотипе исследуемых полиморфизмов генов (их комбинаций) относительные шансы ( OR ) содержания в крови хлороформа в концентрации ≥ Р75 значительно выше, чем у лиц без полиморфизмов ( OR = 29,8; 95 % ДИ: 8,7–102,5). В зависимости от наличия конкретных полиморфизмов отношение шансов варьируется от 8,4 раза (подгруппа № 4 «полиморфизм GSTM1 (моно)») до 59 раз (подгруппа № 19 «GSTT1+EPHX1+CYP2E1») в сравнении с отсутствием в генотипе изучаемых полиморфизмов. Исключение – подгруппы № 7 «полиморфизм CYP2E1 (моно)» и № 18 «полиморфизмы GSTM1+GSTT1+CYP2E1» (концентрация хлороформа < Р75). Наблюдаемые зависимости статистически значимы ( р < 0,05) за исключением

а
Медиана 2SH-7SH Размах Cxi выЬр Kpaiecne точки
б
г
д
Рис. 2. Уровни хлороформа в крови добровольцев группы наблюдения: диаграммы размаха для подгрупп со статистически значимыми различиями: а – с подгруппой № 4; б – с подгруппой № 8; в – с подгруппой № 13;
г – с подгруппой № 14; д – с подгруппой № 19
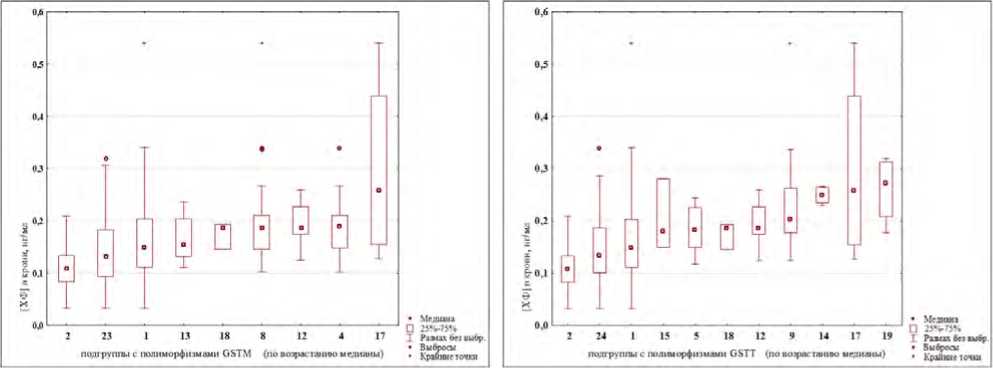
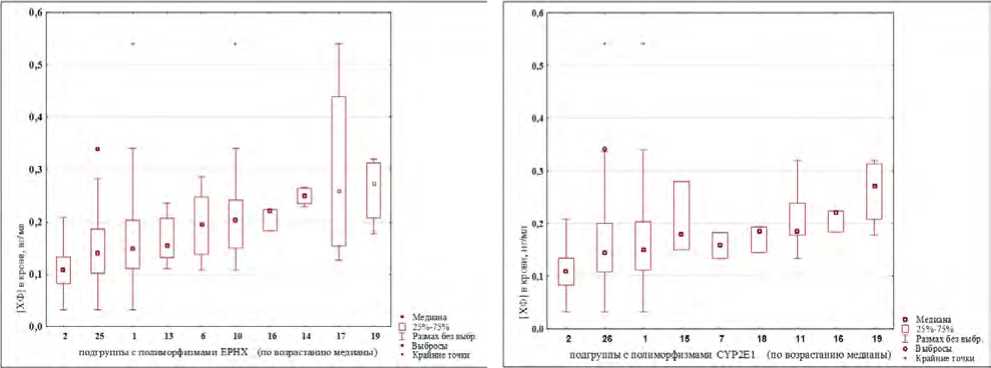
Рис. 3. Уровни хлороформа в крови добровольцев группы наблюдения: диаграммы размаха для подгрупп с наличием / отсутствием полиморфизмов генов CYP2E1, GSTM1, GSTT1, EPHX1 (последовательности подгрупп представлены соответственно возрастанию значений медиан концентраций)
подгруппы № 15. При этом наиболее высокая вероятность того, что в крови экспонированного лица хлороформ содержится в относительно более высокой концентрации (≥ Р75), отмечается при наличии в генотипе полиморфизма гена EPHX1, особенно в сочетании с полиморфизмом гена GSTТ1 (различия по всем комбинациям статистически значимы, р < 0,05) (табл. 4).
Выводы. В группе наблюдения частота генотипов полиморфного локуса C1091T гена CYP2E1 составила 10 %, встречаемости нулевых генотипов GSTM1 и GSTT1 – 36,7 и 23,3 % соответственно, полиморфизма 4-го экзона А415G гена EPHX1 – 15,7 % (мутантных гомозигот GG – 9,3 %, гетерозиготного генотипа AG – 12,7 %).
Наличие в генотипе изученных полиморфизмов (A415G гена EPHX1, C1091T гена CYP2E1, нулевые мутации генов глутатионтрансферазы GSTT1 и GSTM1) ассоциировано с замедлением выведения и биотрансформации хлороформа, его накоплением в организме и повышением равновесных концентраций хлороформа в крови населения, подвергавшегося длительному воздействию побочных продуктов дезин- фекции с водой. При содержании хлороформа в воде в пределах ПДК (0,2 мг/дм3) концентрации хлороформа в крови экспонированного населения составили: Ме = 0,1892 нг/мл [0,1495; 0,2281] у лиц с наличием полиморфизма / полиморфизмов и Ме = 0,1082 нг/мл [0,0822; 0,1341] в группе с нормальными генотипами анализируемых генов (U = 492; Z = 8,53; р < 0,0000001). При этом концентрации хлороформа в крови (по медиане) были выше на 80,4 % и более при наличии в генотипе полиморфизма А415G гена EPHX1, на 68,2 % и более – нулевого аллеля гена GSTT1, на 43,8 % и более – нулевого аллеля гена GSTM1 (различия статистически значимы при р < 0,01).
Для экспонированных лиц с наличием в генотипе исследуемых полиморфизмов генов (их комбинаций) относительные шансы (OR) содержания в крови хлороформа в концентрации ≥ Р75 значительно выше, чем у лиц без полиморфизмов (в 29,8 раза). Присутствие полиморфизма гена EPHX1 существенно увеличивает вероятность содержания хлороформа в крови в более высокой концентрации (≥ Р75, ≥ Р90), особенно в сочетании с полиморфизмом гена GSTТ1.
Таблица 4
Отношения шансов ( OR ) для полиморфных генов и их комбинаций для концентраций > Р75 содержания хлороформа в крови
|
Подгруппа по наличию / отсутствию мутантных аллелей (полиморфизмов) (пояснение) |
OR (95 % ДИ) (для проб с концентрациями хлороформа в крови ≥ Р75 с полиморфизмами к таковым и без полиморфизмов) |
Отношение рисков ( RR ) |
|
3. Есть полиморфизмы |
29,8 (8,7–102,5) |
12,4 |
|
4. Полиморфизм GSTM1 (моно) |
8,4 (21–34,1) |
6,2 |
|
5. Полиморфизм GSTT1 (моно) |
15,7 (2,7–90,8) |
9,2 |
|
6. Полиморфизм EPHX1 (моно) |
19,7 (3,2–119,9) |
10,3 |
|
7. Полиморфизм CYP2E1 (моно) |
0* |
0,0 |
|
8. Полиморфизм GSTM1 (не только моно) |
8,8 (2,4–32,1) |
6,4 |
|
9. Полиморфизм GSTT1 (не только моно) |
18,6 (4,9–70,7) |
10,0 |
|
10. Полиморфизм EPHX1 (не только моно) |
23,6 (6,1–90,8) |
11,3 |
|
11. Полиморфизм CYP2E1 (не только моно) |
13,1 (2,8–62,0) |
8,3 |
|
12. Полиморфизмы GSTM1+GSTT1 |
11,8 (1,9–74,5) |
7,8 |
|
13. Полиморфизмы GSTM1+EPHX1 |
8,4 (1,4–50,1) |
6,2 |
|
14. Полиморфизмы GSTT1+EPHX1 |
–** |
20,7 |
|
15. Полиморфизмы GSTT1+CYP2E1 |
9,8 (0,7–141,4)*** |
6,9 |
|
16. Полиморфизмы EPHX1+CYP2E1 |
39,3 (2,7–565,8) |
13,8 |
|
17. Полиморфизмы GSTM1+GSTT1+EPHX1 |
19,7 (2,0–191,8) |
10,3 |
|
18. Полиморфизмы GSTM1+GSTT1+CYP2E1 |
0* |
0,0 |
|
19. Полиморфизмы GSTT1+EPHX1+CYP2E1 |
59,0 (4,6–750,5) |
15,5 |
|
20. Полиморфизмы более 1 гена |
16,9 (4,5–63,1) |
9,5 |
|
21. Полиморфизмы двух генов |
51,1 (10,8–241,5) |
14,9 |
|
22. Полиморфизмы трех генов |
24,6 (4,3–141,9) |
11,5 |
П р и м е ч а н и е : * – в подгруппе с наличием данных полиморфизмов все пробы крови содержали хлороформ ниже Р75, ** – ОШ невозможно рассчитать, поскольку в подгруппе с наличием полиморфизмов все пробы содержат хлороформ > Р75, *** - установленные различия недостоверны, p > 0,05.
Установленные особенности позволяют рассматривать гетеро- и гомозиготные полиморфные генотипы AG/GG по гену EPHX1, CT/ТТ по гену CYP2E1, а также наличие нулевого аллеля в генах GSTT1 и GSTM1 в качестве факторов генетической предрасположенности к накоплению хлороформа в организме, что способствует повышению вероятности реализации рисков здоровью, ассоциированных с хроническим воздействием данного побочного продукта дезинфекции. Учитывая вышеизложенное, указанные полиморфизмы могут выступать в качестве биомаркеров индивидуальной чувствительности при оценке подверженности населения воздействию хлороформа, поступающего в организм с водой централизованных систем хозяйственнопитьевого водоснабжения, и применяться для совершенствования мер профилактики.
В качестве наиболее информативных биомаркеров чувствительности при оценке рисков, ассоциированных с воздействием хлороформа на уровнях, не
превышающих ПДК в воде, предлагается использовать полиморфизм A415G гена EPHX1 и делецию гена глутатионтрансферазы GSTT1, а также их комбинации, в том числе с делецией гена GSTM1 и / или полиморфизмом C1091T гена CYP2E1.
Перспективным направлением дальнейших исследований представляется изучение ассоциаций между воздействием тригалометанов и бременем патогенетически обусловленных заболеваний с учетом генетической восприимчивости популяции.
Финансирование. Работа выполнялась в рамках задания 01.01. «Разработать метод гигиенической оценки летучих химических веществ в питьевой воде» подпрограммы «Безопасность среды обитания человека» ГНТП «Научно-техническое обеспечение качества и доступности медицинских услуг», 2021–2025 годы (рег. № НИОКР 20190177).
Список литературы Полиморфные варианты генов ферментов системы детоксикации ксенобиотиков CYP2Е1, GSTM1, GSTT1, EPHX1 как биомаркеры чувствительности к экспозиции побочными продуктами водоподготовки (на примере хлороформа)
- Guidelines for Drinking-water Quality, 4th ed. with adds. - Geneva: World Health Organization, 2017. - 564 р.
- Sharma V.K., Zboril R., McDonald T.J. Formation and toxicity of brominated disinfection byproducts during chlorination and chloramination of water: a review // J. Environ. Sci. Health B. - 2014. - Vol. 49, № 3. - P. 212-228. DOI: 10.1080/03601234.2014.858576
- Егорова Н.А., Букшук А.А., Красовский Г.Н. Гигиеническая оценка продуктов хлорирования питьевой воды с учетом множественности путей поступления в организм // Гигиена и санитария. - 2013. - Т. 92, № 2. - С. 18-24.
- EPA/600/R-06/087. Exposures and internal doses of trihalomethanes in humans: multi-route contributions from drinking water [Электронный ресурс]. - URL: http://nepis.epa.gov/Adobe/PDFApdf (дата обращения: 15.01.2018).
- Kujlu R., Mahdavianpour M., Ghanbari F. Multi-route human health risk assessment from trihalomethanes in drinking and non-drinking water in Abadan, Iran // Environmental Science and Pollution Research. - 2020. - Vol. 27. - P. 42621-42630.
- Health impacts of long-term exposure to disinfection by-products in drinking water in Europe: HIWATE / M.J. Nieu-wenhuijsen, R. Smith, S. Golfinopoulos, N. Best, J. Bennett, G. Aggazzotti, E. Righi, G. Fantuzzi [et al.] // J. Water Health. -2009. - Vol. 7, № 2. - P. 185-207. DOI: 10.2166/wh.2009.073
- Occurrence, genotoxicity, and carcinogenicity of regulated and emerging disinfection byproducts in drinking water: a review and roadmap for research / S.D. Richardson, M.J. Plewa, E.D. Wagner, R. Schoeny, D.M. Demarini // Mutat. Res. -2007. - Vol. 636, № 1-3. - P. 178-242. DOI: 10.1016/j.mrrev.2007.09.001
- К вопросу об образовании побочных продуктов дезинфекции питьевой воды (регламентируемых и эмерджентных), их генотоксических и канцерогенных свойствах: обзор проблемы и направления дальнейших исследований / Е.В. Дроздова, В.В. Бурая, В.В. Гирина, Т.З. Суровец, А.В. Фираго // Здоровье и окружающая среда. - 2016. - № 26. - С. 12-16.
- Tellez Tovar S.S., Rodriguez Susa M. Cancer risk assessment from exposure to trihalomethanes in showers by inhalation // Environ. Res. - 2021. - Vol. 196. - P. 110401. DOI: 10.1016/j.envres.2020.110401
- Trihalomethanes in Drinking Water and Bladder Cancer Burden in the European Union / I. Evlampidou, L. FontRibera, D. Rojas-Rueda, E. Gracia-Lavedan, N. Costet, N. Pearce, P. Vineis, J.J.K. Jaakkola [et al.] // Environ. Health Perspect. -2020. - Vol. 128, № 1. - P. 17001. DOI: 10.1289/EHP4495
- Colorectal cancer and long-term exposure to trihalomethanes in drinking water: a multicenter case-control study in Spain and Italy / C.M. Villanueva, E. Gracia-Lavedan, C. Bosetti, E. Righi [et al.] // Environ. Health Perspect. - 2017. -Vol. 125, № 1. - Р. 56-65. DOI: 10.1289/EHP155
- Экспериментальные модели хронической патологии животных для оценки рисков здоровью чувствительных групп населения / Е.В. Дроздова, С.И. Сычик, В.А. Грынчак, С.Н. Рябцева // Анализ риска здоровью. - 2022. -№ 2. - С. 185-195. DOI: 10.21668/health.risk/2022.2.17
- Household exposures to drinking water disinfection by-products: whole blood trihalomethane levels / L.C. Backer, D.L. Ashley, M.A. Bonin, F.L. Cardinali, S.M. Kieszak, J.V. Wooten // J. Expo. Anal. Environ. Epidemiol. - 2000. - Vol. 10, № 4. - P. 321-326. DOI: 10.1038/sj.jea.7500098
- Опыт установления и доказывания вреда здоровью населения вследствие потребления питьевой воды, содержащей продукты гиперхлорирования / Н.В. Зайцева, И.В. Май, С.В. Клейн, Э.В. Седусова // Здоровье населения и среда обитания - ЗНиСО. - 2015. - Т. 273, № 12. - С. 16-18.
- Биохимические маркерные показатели негативных эффектов у детей при воздействии хлорорганических соединений с питьевой водой / М.А. Землянова, О.В. Пустовалова, Д.Л. Мазунина, А.С. Сбоев // Гигиена и санитария. -2016. - Т. 95, № 1. - С. 97-101. DOI: 10.18821/0016-9900-2016-95-1-97-101
- Четверкина К.В. К установлению реперного уровня содержания хлороформа в крови детского населения // Анализ риска здоровью. - 2018. - № 3. - С. 85-93. DOI: 10.21668/health.risk/2018.3.09
- Human exposure assessment for DBPs: factors influencing blood trihalomethane levels / B.C. Blount, L.L. Aylward, J. Lakind, L.S. Backer, S.M. Hays // Encyclopedia ofEnvironmental Health. - 2011. - Vol. 3. - P. 100-107. DOI: 10.1016/B978-0-444-52272-6.00103-3
- Changes in blood trihalomethane concentrations resulting from differences in water quality and water use activities / D.L. Ashley, B.C. Blount, P.C. Singer, E. Depaz, C. Wilkes, S. Gordon, C. Lyu, J. Masters // Arch. Environ. Occup. Health. -2005. - Vol. 60, № 1. - Р. 7-15. DOI: 10.3200/AEOH.60.1.7-15
- Influence of tap water quality and household water use activities on indoor air and internal dose levels of triha-lomethanes / J.R. Nuckols, D.L. Ashley, C. Lyu, S.M. Gordon, A.F. Hinckley, P. Singer // Environ. Health Perspect. - 2005. -Vol. 113, № 7. - Р. 863-870. DOI: 10.1289/ehp.7141
- Exogenous and Endogenous Determinants of Blood Trihalomethane Levels after Showering / L.C. Backer, Q. Lan, B.C. Blount, J.R. Nuckols [et al.] // Environ. Health Perpect. - 2008. - Vol. 116, № 1. - Р. 57-63. DOI: 10.1289/ehp.10049
- Predictors of blood trihalomethane concentrations in NHANES 1999-2006 / А.М. Riederer, R. Dhingra, B.C. Blount, K. Steenland // Environ. Health Perspect. - 2014. - Vol. 122, № 7. - P. 695-702. DOI: 10.1289/ehp.1306499
- Xenobiotic sensor- and metabolism-related gene variants in environmental sensitivity-related illnesses: a survey on the Italian population / D. Caccamo, E. Cesareo, S. Mariani, D. Raskovic [et al.] // Oxid. Med. Cell. Longev. - 2013. -Vol. 2013. - P. 831969. DOI: 10.1155/2013/831969
- FOXE1 polymorphisms and chronic exposure to nitrates in drinking water cause metabolic dysfunction, thyroid abnormalities, and genotoxic damage in women / D.D. Gandarilla-Esparza, E.Y. Calleros-Rincón, H.M. Macias, M.F. González-Delgado [et al.] // Genet. Mol. Biol. - 2021. - Vol. 44, № 3. - P. e20210020. DOI: 10.1590/1678-4685-GMB-2021-0020
- Markers of genetic susceptibility in human environmental hygiene and toxicology: the role of selected CYP, NAT and GST genes / R. Thier, T. Brüning, P.H. Roos, H.-P. Rihs [et al.] // Int. J. Hyg. Environ. Health. - 2003. - Vol. 206, № 3. -Р. 149-171. DOI: 10.1078/1438-4639-00209
- Autrup H. Genetic polymorphysms in human xenobiotica metabolizing enzymes as suscectibility factors in toxic response // Mutat. Res. - 2000. - Vol. 464, № 1. - P. 65-76. DOI: 10.1016/s1383-5718(99)00167-9
- DNA methylation levels and long-term trihalomethane exposure in drinking water: an epigenome-wide association study / L.A. Salas, M. Bustamante, J.R. Gonzalez, E. Gracia-Lavedan [et al.] // Epigenetics. - 2015. - Vol. 10, № 7. - P. 650-661. DOI: 10.1080/15592294.2015.1057672
- Drinking Water Disinfection By-products, Genetic Polymorphisms, and Birth Outcomes in a European Mother-Child Cohort Study / M. Kogevinas, M. Bustamante, E. Gracia-Lavedán, F. Ballester [et al.] // Epidemiology. - 2016. - Vol. 27, № 6. - P. 903-911. DOI: 10.1097/EDE.0000000000000544
- Polymorphisms in GSTT1, GSTZ1, and CYP2E1, Disinfection By-products, and Risk of Bladder Cancer in Spain / K.P. Cantor, C.M. Villanueva, D.T. Silverman, J.D. Figueroa [et al.] // Environ. Health Perspect. - 2010. - Vol. 118, № 11. -P. 1545-1550. DOI: 10.1289/ehp.1002206
- Infante-Rivard C. Drinking water contaminants, gene polymorphisms, and fetal growth // Environ. Health Perspect. -2004. - Vol. 112, № 11. - P. 1213-1216. DOI: 10.1289/ehp.7003
- Effect modification of CYP2E1 and GSTZ1 genetic polymorphisms on associations between prenatal disinfection by-products exposure and birth outcomes / B. Zhou, P. Yang, Y.-J. Gong, Q. Zeng, W.-Q. Lu, X.-P. Miao // Environ. Pollut. -2018. - Vol. 243, Pt B. - P. 1126-1133. DOI: 10.1016/j.envpol.2018.09.083
- Prenatal exposure to drinking-water chlorination by-products, cytochrome P450 gene polymorphisms and small-for-gestational-age neonates / S.G. Bonou, P. Levallois, Y. Giguere, M. Rodriguez, A. Bureau // Reprod. Toxicol. - 2017. -Vol. 73. - P. 75-86. DOI: 10.1016/j.reprotox.2017.07.019
- Interactions between CYP2E1, GSTZ1 and GSTT1 polymorphisms and exposure to drinking water trihalomethanes and their association with semen quality / P. Yang, Q. Zeng, W.-C. Cao, Y.-X. Wang [et al.] // Environ. Res. - 2016. -Vol. 147. - P. 445-452. DOI: 10.1016/j.envres.2016.03.009
- Metabolic gene polymorphism frequencies in control populations / S. Garte, L. Gaspari, A.K. Alexandrie, C. Am-brosone [et al.] // Cancer Epidemiol. Biomarkers Prev. - 2001. - Vol. 10, № 12. - Р. 1239-1248.
- Содержание хлороформа в крови населения как биомаркер экспозиции побочными продуктами дезинфекции питьевой воды / Е.В. Дроздова, С.И. Сычик, В.Э. Сяхович, Е.Н. Походня [и др.] // Медицинский журнал. - 2023. -Т. 83, № 1. - С. 23-32. DOI: 10.51922/1818-426X.2023.1.23
