Построение спектральной плотности решения линейного стохастического дифференциального уравнения в частных производных с постоянными запаздываниями

Автор: Полосков И.Е.
Журнал: Вестник Пермского университета. Математика. Механика. Информатика @vestnik-psu-mmi
Рубрика: Механика. Математическое моделирование
Статья в выпуске: 1 (40), 2018 года.
Бесплатный доступ
Данная работа посвящена распространению схемы Гийюзика (S.Guillouzic), предложенной для вычисления спектральной плотности решения линейного стохастического дифференциального уравнения первого порядка с постоянными коэффициентами и запаздыванием, на новое семейство уравнений - стохастические эволюционные дифференциальные уравнения в частных производных с несколькими постоянными запаздываниями. Задача исследования состояла в построении спектральной плотности стационарного случайного поля - решения гиперболического уравнения с постоянными коэффициентами и случайным входом.
Спектральная плотность, стохастическое дифференциальное уравнение в частных производных, запаздывание, стационарное случайное поле, условие существования спектральной плотности
Короткий адрес: https://sciup.org/147245354
IDR: 147245354 | УДК: 519.2 | DOI: 10.17072/1993-0550-2018-1-36-45
Derivation of the spectral density function for solution of linear stochastic partial differential equation with constant delays
This paper is devoted to extension of the scheme of S. Guillouzic, proposed to calculate the spectral density functions for solutions of linear stochastic differential equations of the first order with constant coefficients and one fixed delay, to a new family of equations, i.e. stochastic evolutionary partial differential equations with few constant delays. The aim of our study was to construct the spectral density function of a stationary random field as the solution of hyperbolic equation with fixed coefficients and with random input.
Текст научной статьи Построение спектральной плотности решения линейного стохастического дифференциального уравнения в частных производных с постоянными запаздываниями
Предисловие
Необходимость математического моделирования роста растений связана с земледелием двояко: с одной стороны – для обеспечения текущих потребностей агротехнологии выращивания культурных растений (продуктов питания), с другой стороны – для селекционной работы по выведению новых, более продуктивных и более приспособленных к внешним условиям сортов растений. Понятно, что широта (в отличие от узкоспециализированно-сти) применения матмоделей обеспечивается их простотой и простотой интерпретации их параметров. Ниже описаны достаточно простые модели роста однолетних растений, использующие аппарат классической теории автоматического управления.
1. "Дифференциальный" подход к моделям
Исторически сложилось так, что математические модели роста растений рассматривались как модели скорости роста биомассы, например: "для свободно растущего рас-
тения две функции возраста Аj(t) – количество средств существования j-го типа и B(t) – скорость роста биомассы свободно растущего растения возраста t" [1, с. 715]1, где B(t) может быть вектором, учитывающим рост биомассы отдельных органов растения (корней, листьев, стеблей, плодов и т. п.). В такой "дифференциальной" форме модель представляла собой систему дифференциальных уравнений, и если в модель вводилось управление (для получения максимальных урожаев), то оно, соответственно принципу максимума Понтрягина, давало решение на границах допустимой области управлений: максимум допустимой освещенности, максимум допустимого полива, максимум допустимых доз минеральных удобрений2 и т. п. Однако не всеми параметрами можно управлять, особенно это касается климатических параметров, и задача селекции – получение сортов, продуктивных как раз в неоптимальных условиях роста растения (эта общая задача практически неизменна за всю историю селекции [15]).
Проблемы получения математических моделей роста растений отмечались как в 1980–1990-е гг.: "В настоящее время <1990-е гг.> одним из главных препятствий на пути целенаправленного создания агрохимически эффективных сортов является отсутствие моделей как концептуального уровня, так и основанных на экспериментальных данных" [3, с. 302], – так и в недавнем прошлом: "необходимо отметить заметный скепсис ученых-физиологов по поводу возможностей современных математических дисциплин при моделировании продукционных, в том числе и ростовых процессов" [6, с. 103].
Одним из препятствий построения математических моделей роста растений является как раз "дифференциальная" форма модели, затрудняющая интерпретацию и идентификацию параметров модели. "Эмпирические параметры <дифференциальных> уравнений также несут на себе отпечаток случайности и не могут с большой степенью достоверности использоваться для прогнозирования темпов роста растений и формирования урожая" [6, с. 103].
Пример одной из "дифференциальных" моделей роста растений, многократно проверенный на экспериментальных данных приведен ниже.
2. Пример модели и данных
Информационная структура математической модели роста картофеля [11] показана на рис. 1.
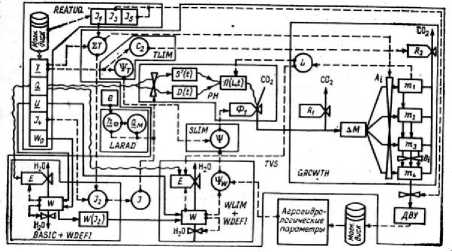
Рис. 1. Структура модели роста картофеля, по [11, с. 86]
Однако ключевым для модели является представление данных в этой модели, пример эмпирических данных и модельного приближения приведен на рис. 2 в виде относитель- ной биомассы органов растения, приведенной в каждый момент роста к 1, показывающей в "дифференциальном" виде скорость роста от-3
дельных органов растения.
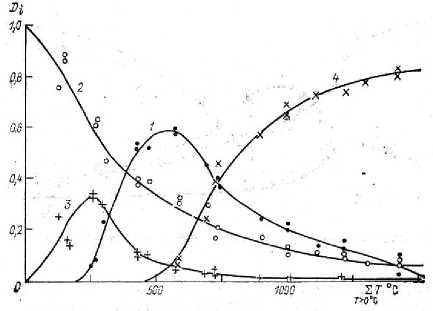
Рис. 2. Данные и результат моделирования для картофеля сорта Белорусский ранний, относительные биомассы: 1 – листья, 2 – стебли, 3 – корни, 4 – клубни, по [11, с. 86] ( по горизонтали – сумма средних температур за день, больших 0 ° С)
Значительная нелинейность функций роста, представленных на рис. 2, затрудняет их сопоставление с действительными процессами роста растения. Если же данные рис. 2 проинтегрировать и привести к виду перенормированному к 1 относительно накопленной к концу роста (сбору урожая) биомассе, то получается простая картина (см. рис. 3, 4).
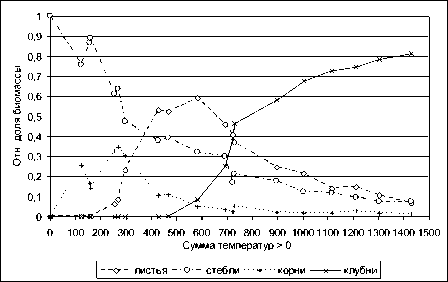
Рис. 3. Экспериментальные данные из рис. 2, [11, с. 86], "дифференциальная" форма
3 О функциях роста в аналогичной форме см. также [12, с. 67].
4. Модель растения как конечная система
5. Модель роста как конечный набор звеньев
Более современные модели, хотя и отображали результаты в "интегральной" форме [8, с. 330, рис. 7.4], но для моделей использовали функции роста в дифференциальной форме [8, с. 299–300], см. также п. 2.
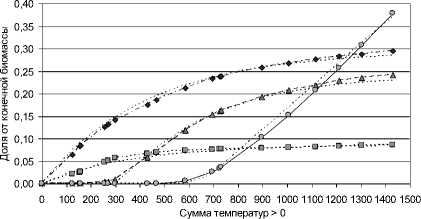
-4- листья -■■♦■- стебли ------- корни клубни стебли_м корни_м листья_м клубни_м
Рис. 4. Проинтегрированные данные из рис. 2, 3 , "интегральная" форма, относительно конечной биомассы, принятой за 1
Кривые на рис. 4 содержательно понятны относительно предметной области (накопления органами растением биомассы), и, кроме того, по теории автоматического управления кривые на рис. 4 хорошо приближаемы функциями реакции на единичное воздействие типовых динамических звеньев (1–2-го порядка) и их комбинаций. О том, почему содержательно допустимо такое приближение, сказано ниже.
Хотя в фундаментальном смысле геномом (и вообще наследственностью) не определяются полностью свойства живого организма (в частности свойство быть живым)4, для прикладных целей, в том плане, что имеется дело с живыми организмами в благоприятной для них среде, с определенным набором протекающих в них и между ними и окружающей средой процессов, в первом приближении модель растения выглядит как конечная система, описываемая приближенно некоторой совокупностью линейных звеньев порядка не выше 2. В этой модели генотип и окружающая среда рассматриваются неразрывно, соответственно высказанным еще в середине XX в., и многократно проверенным к нач. XXI положениям биологической нау-ки5. Таким образом, взгляд на рост растения как на определенный и генотипом и средой некий переходный процесс от начального до конечного состояния (с перенормировкой по конечному состоянию) позволяет усматривать в этом процессе некие достаточно простые закономерности.
В простой модели роста растения его рост рассматривается как реакция на единичное воздействие (функцию 1(t)), соответствующее моменту времени t0 – посеву семени в почву (при благоприятных для роста внешних условиях). Тогда рост органов растения как реакция на это воздействие есть комбинация функций типовых звеньев: запаздывающего звена, апериодического звена, интегрирующего звена (см. [9], [4]). Тогда для "интегральных" данных приведенных на рис. 4 функции модели роста картофеля (см. рис. 2–4) таковы, как указано в таблице: листья – апериодичес-ское звено с запаздыванием; стебли – апериодическое звено; корни – апериодическое звено; клубни – интегрирующее звено c запаздыванием. (На рис. 2–4 физическое время заменено на "биологическое" – сумму температур, что не меняет вида закономерностей; условное время обозначено за t).
Кроме того, имеется общий коэффициент модели k0, переводящий относительные величины рис. 4 и таблицы (h1–h4) в конечную (на сбор урожая) биомассу в ц/га.
По рис. 4 и таблице легко видеть, что запаздывание роста клубней примерно равно двойному запаздыванию роста листьев, относительно начала роста, τ4≈2τ1. Постоянные ни, своего развития и определённо реагировать на те или иные условия" [5]. К концу XX – нач. XXI в. единство в жизни растения генотипа и условий среды было опытно подтверждено, вплоть до влияния среды на генотип, а именно была "показана способность растений пшеницы (а может быть и всего класса эукариот) к переопределению генетических формул сорта под влиянием обеспеченности элементами питания и другими факторами среды, как <…> механизм неспецифической природы" [3, с. 296], необходимый для сохранения вида. И, ближе к современности, установлено, что "изменение определенных условий окружающей среды приводит к изменению локализации <в геноме> QTL (хозяйственно-ценных признаков), вовлеченных в реализацию некоторых признаков <у яровой мягкой пшеницы>" [13, с. 65].
времени (T 1 –T 3 ) роста служебных органов (не урожая) приближенно равны.
Функции модели для отдельных органов
|
Орган |
Формула реакции на 1(t) |
Параметры* |
|
Листья |
h 1 (t)= 1( τ 1 ) ⋅ k 1 (1–exp(–(t– τ 1 )/T 1 ) |
k 1 =0,229; Т 1 =370; τ 1 =320 |
|
Стебли |
h 2 (t)= 1(0) ⋅ k 2 (1–exp(–t/T 2 ) |
k 2 =0,284; Т 2 =420 |
|
Корни |
h 3 (t)= 1(0) ⋅ k 3 (1–exp(–t/T 3 ) |
k 3 =0,084; Т 3 =370 |
|
Клубни |
h 4 (t)= 1( τ 4 ) ⋅ k 4 (t– τ 4 )/T 4 |
k 4 =0,371; Т 4 =2100; τ 4 =650 |
*k i – безразмерны, Т i , τ i – в условных единицах биологического времени, для данных рис. 2–4 – сумма дневных температур, больших 0
Какие параметры столбца 3 таблицы, и в какой мере определяются генотипом и условиями роста – подлежит отдельному опытному определению для конкретных сортов рас-тений6.
Модель позволяет определить особенности роста урожая, в отличие от роста других органов: рост урожая описывается интегральным звеном с запаздыванием.
6. Примеры приложения модели
Примеры интерпретации модели приведены по оцифрованным интегрированным с перенормировкой данным. Для ячменя получается следующая картина (рис. 5): – "дифференциальные" данные (из [10]), понимание затруднено; рис. 6 – проинтегрированные данные, интерпретация аналогична таковой в п. 5.
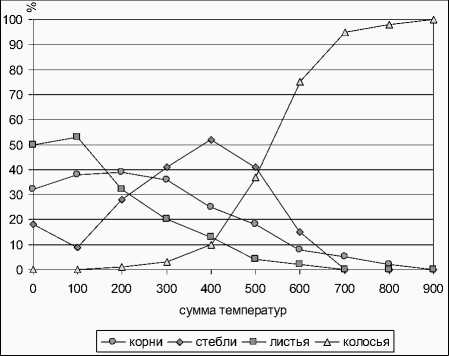
Рис. 5. Функции текущей приведенной к 1 биомассы для ячменя, по [10]
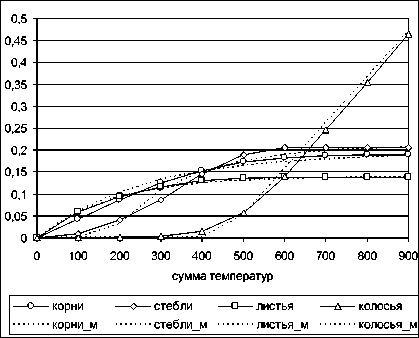
Рис. 6. Функции модели для ячменя, по рис. 5, см. п. 5
Для ячменя, как и для картофеля, по модели (рис. 6) легко различимы функции роста служебных органов и урожая (колосьев); для пшеницы получается несколько иной результат, – к функции роста урожая относится не только рост колосьев, но и стеблей (т. е. пшеница, кроме зерна, на урожай, как на результат интегрального роста биомассы, дает и солому), см. рис. 7.
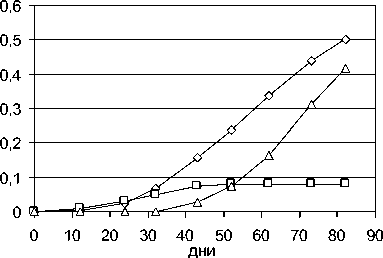
стебли листья колосья
Рис. 6. Приближенные функции роста для пшеницы, к итоговой биомассе, по проинтегрированными и перенормированным данным из [10]
Заключение
Таким образом, с использованием интегральных данных, приведенных к общей биомассе растения на конец роста, посредством стандартных средств теории автоматического управления (функций реакций основных передаточных звеньев на единичное воздействие), построены простые модели роста культурных растений, допускающие ясную качественную интерпретацию, и требующие дальнейшего определения количественных параметров.
Список литературы Построение спектральной плотности решения линейного стохастического дифференциального уравнения в частных производных с постоянными запаздываниями
- Беллман Р., Кук К. Дифференциально-разностные уравнения. М.: Мир, 1967. 548 с
- Хейл Дэю. Теория функционально-дифференциальных уравнений. М.: Мир, 1984. 421 с
- Рубаник В.П. Колебания сложных квазилинейных систем с запаздыванием. Мн.: Изд-во "Университетское", 1985. 143 с.
- Царьков Е. Ф. Случайные возмущения дифференциально-функциональных уравнений. Рига: Зинатне, 1989. 421 с.
- Элъсголъц Л.Э., Норкин С.Б. Введение в теорию дифференциальных уравнений с отклоняющимся аргументом. М.: Наука, 1971. 296 с.
- Boukas El-K., Liu Zi-K. Deterministic and stochastic time delay systems. Boston: Birkhauser, 2002. XVI, 423 p.
- Driver R.D. Ordinary and delay differential equations. New York, Heidelberg, Berlin: Springer, 1977. IX, 501 p.
- Маланин В.В., Полосков И.Е. Методы и практика анализа случайных процессов в динамических системах: учеб. пособие. Ижевск: РХД, 2005. 296 с.
- Полосков И.Е. Стохастический анализ динамических систем [Электронный ресурс]: монография. Пермь: Изд-во Перм. ун-та, 2016. 772 с.
- Garrido T.G. Existence and uniqueness of solutions for non-linear stochastic partial differential equations // Collectanea Mathematica. 1991. Vol. 42, № 1. P. 51-74.
- Chow P.-L. Stochastic partial differential equations. Boca Raton (FL): Chapman & Hall / CRC, 2015. XII, 314 p.
- Mandrekar V.S., Gawarecki L. Stochastic analysis for Gaussian random processes and fields with applications. Boca Raton (FL): Chapman & Hall / CRC, 2016. XXI, [1], 179 p.
- Caraballo Т., Real J., Taniguchi T. The exponential stability of neutral stochastic delay partial differential equations // Discrete and Continuous Dynamical Systems. Ser. A. 2007. Vol. 18, № 2-3. P. 295-313.
- Chang M.-H. Weak infinitesimal generator for a stochastic partial differential equation with time delay // Journal of Applied Mathematics and Stochastic Analysis. 1995. Vol.8, № 2. P. 115-138.
- Galtier M. Touboul J. On an explicit representation of the solution of linear stochastic partial differential equations with delays // Comptes Rendus Mathematique. 2012. Vol. 350, № 3-4. P. 167-172.
- Jahanipur R. Stochastic functional evolution equations with monotone nonlinearity: Existence and stability of the mild solutions // Journal of Differential Equations. 2010. Vol. 248, № 5. P. 12301255.
- Liu K., Truman A. Lyapunov function approaches and asymptotic stability of stochastic evolution equations in Hilbert spaces - A survey of recent developments // Stochastic partial differential equations and applications / G. Da Prato, L.Tubaro (eds.). New York: Marcel Dekker, 2002. P. 337-372.
- Luo Q., Deng F., Bao J. Zhao B. Sliding mode control of a class of Ito type distributed parameter systems with delay // Acta Mathe-matica Scientia. 2007. Vol. 27B, № 1. P. 67-76.
- Pan L., Zhong S. Dynamic analysis of stochastic reaction-diffusion Cohen-Grossberg neural networks with delays // Advances in Difference Equations. 2009. Vol. 2009. Article ID 410823. 18 p.
- Frank T.D., Beek P.J. Stationary solutions of linear stochastic delay differential equations: Applications to biological systems // Physical Review. 2001. Vol. E64, № 2. P. 1:021917. 12 p.
- Frank T.D. Multivariate Markov processes for stochastic systems with delays: Application to the stochastic Gompertz model with delay // Physical Review. 2002. V0I.E66, № 1. P. 1:011914. 8 p.
- Frank T.D. Stationary distributions of stochastic processes described by a linear neutral delay differential equation // Journal of Physics A: Mathematical and General. 2005. Vol. 38, № 28. P. L485-L490.
- Frank T.D. Delay Fokker-Planck equations, Novikov's theorem, and Boltzmann distributions as small delay approximations // Physical Review. 2005. Vol.E72, № 1. P.l:011112. 8 p.
- Guillouzic S., L'Heureux I., Longtin A. Small delay approximation of stochastic differential delay equations // Physical Review. 1999. Vol.E59, № 4. P. 3970-3982.
- Guillouzic S., L'Heureux I., Longtin A. Rate processes in a stochastically driven delayed overdamped // Physical Review. 2000. V0I.E6I, № 5. P. 4906-4914.
- Guillouzic S. Fokker-Planck approach to stochastic delay differential equations. Thesis.. Doctor of Philosophy. Ottawa: University of Ottawa, 2000. 200 p.
- Kuchler U., Mensch B. Langevin's stochastic differential equation extended by a time-delayed term // Stochastics and Stochastic Reports. 1992. Vol.40, № 1-2. P. 23-42.
- VanMarcke E. Random fields: Analysis and synthesis. Cambridge: MIT Press, 1983. 382 p. (Web Edition by Rare Book Services, Princeton (NJ): Princeton University Press, 1998).
