Продуктивные качества, переваримость кормов и кишечный микробиом у цыплят-бройлеров (Gallus gallus L.) при добавлении в рацион нативных и экструдированных углеводных компонентов
 при добавлении в рацион нативных и экструдированных углеводных компонентов")
Автор: Холодилина Т.Н., Яушева Е.В., Рязанцева К.В., Сизова Е.А., Нечитайло К.С.
Журнал: Сельскохозяйственная биология @agrobiology
Рубрика: Технологии кормления
Статья в выпуске: 2 т.59, 2024 года.
Бесплатный доступ
Экструзия как метод термической и механической обработки, сочетающий высокие температуры и давление, повышает пищевую ценность кормовых ингредиентов и широко используется для улучшения функциональных свойств пищевых продуктов. Однако неясно, как рационы, включающие экструдаты, влияют на микробиоту кишечника и физиологию питания цыплят-бройлеров. В настоящей работе впервые установлено, что замена до 10 % пшеницы на отруби и кукурузу не ухудшает продуктивность птицы. Выявлено снижение усвоения жира при экструзии отрубей и повышение этого показателя при обработке кукурузы. Показано, что микробиом слепой кишки по-разному формируется при различных соотношениях растворимых и нерастворимых углеводов. Наша цель заключалась в оценке влияния замены зерновой части рациона цыплят-бройлеров на нативные и экструдированные пшеничные и кукурузные отруби на показатели роста, переваримость и состав кишечного микробиома. Исследования проводили в 2023 году в виварии ФНЦ биологических систем и агротехнологий РАН. Цыплят-бройлеров (Gallus gallus L.) кросса Arbor Acres разделили на четыре опытных группы (по n = 5) методом пар-аналогов. Птицу кормили полнорационными комбикормами согласно рекомендациям Всероссийского научно-исследовательского и технологического института птицеводства (ВНИТИП). Начиная с 21-х сут, 10 % (100 г/кг рациона) зерна пшеницы в рационе цыплят I группы заменяли на пшеничные отруби (О), II группы - на экструдат отрубей (ЭО), III группы - на кукурузу (К), IV группы - на экструдат кукурузы (ЭК). Содержание сырой клетчатки в исследуемых компонентах было следующим: О - 7,7 %, ЭО - 2,7 %, К - 4,29 %, ЭК - 1,67 %. Прирост живой массы учитывали по результатам еженедельного взвешивания (21-42-е сут жизни). Исследовали химический состав рационов и помета. Количество сухого вещества определяли по ГОСТ 31640-2012 (M., 2012), массовую долю сырого жира - по ГОСТ 13496.15-2016 (M., 2016), сырого протеина - по ГОСТ 13496.4-93 (M., 2011), сырой клетчатки - по ГОСТ 31675-2012 (M., 2014). После окончания эксперимента на 42-е сут проводили убой птицы. Органы пищеварения (мышечный и железистый желудки, кишечник) извлекали, освобождали от содержимого и взвешивали. Отбор содержимого слепой кишки проводили во время убоя. Проводили подготовку библиотек ДНК, секвенирование и биоинформационную обработку. Визуализацию результатов биоинформатической обработки и статистический анализ осуществляли с помощью MicrobiomeAnalyst (https://www.microbiomeanalyst.ca/). Полученные OTU после фильтрации и определения таксономической принадлежности использовали для расчета показателей альфа-разнообразия (индекс Chao1, индекс Фишера, индекс биоразнообразия Шеннона, оценка охвата на основе численности ACE, индекс разнообразия Симпсона; статистический метод ANOVA) и бета-разнообразия (метод ординации NMDS, дистанционный метод, индекс Брея-Кертиса; статистический метод PERMANOVA). На 14-е сут после начала эксперимента наблюдалась тенденция к увеличению живой массы в группах, получавших экструдированные продукты, - на 8,5 % для ЭО и 9,9 % для ЭК в сравнении с аналогичными кормами без экструзии. К концу учетного периода разница для тех же групп по живой массе составила соответственно 8,3 и 10,4 %, значимых различий не наблюдалось. Внесение в рационы экструдированной кукурузы приводило к статистически значимому увеличению переваримости сырого жира (СЖ): этот показатель был на 5,1 % (р ≤ 0,05) выше в сравнении с ЭО и на 6,4 % (р ≤ 0,05) при сравнении с К. Экструдированные отруби в рационе цыплят-бройлеров снижали переваримость СЖ на 3,5 % при сравнении с I группой. При оценке переваримости сырого протеина (СП) отмечали обратный эффект: происходило статистически значимое снижение в III и IV группах соответственно на 6,6 % (р ≤ 0,05) и 13,2 % (р ≤ 0,05) по сравнению с I группой. Остальные показатели усвоения питательных веществ не имели достоверных различий. На развитие органов пищеварения влияло более высокое содержание клетчатки в нативных компонентах отрубей и кукурузы. Масса мышечного желудка в I группе была на 7,02 % (p ≤ 0,001) выше, чем у птиц, которым скармливали ЭО, в III группе - на 22,04 % (p ≤ 0,05) выше, чем при скармливании ЭК. Масса кишечника снижалась в IV группе на 22,12 % (p function show_abstract() { $('#abstract1').hide(); $('#abstract2').show(); $('#abstract_expand').hide(); }
Цыплята, кормление, экструзия, кукуруза, пшеничные отруби, продуктивность, кишечный микробиом
Короткий адрес: https://sciup.org/142242445
IDR: 142242445 | УДК: 636.52/.58:636.085.8:591.132.6:577.2 | DOI: 10.15389/agrobiology.2024.2.274rus
Productive performance, feed digestibility and gut microbiome of broiler chickens (Gallus gallus L.) fed diets with native and extruded carbohydrate containing ingredients
Extrusion, as a thermal and mechanical processing method combining high temperatures and pressure, increases the nutritional value of feed ingredients and it is widely used to improve the functional properties of food products. However, it is unclear how diets containing extrudates interact with the intestinal microbiota and influence the nutritional physiology of broiler chickens. In this work, it was established for the first time that replacing up to 10 % of wheat with bran and corn does not affect poultry productivity. A decrease in fat absorption during bran extrusion and an increase in this indicator during corn processing were revealed. It has been shown that the cecal microbiome is formed differently at different ratios of soluble and insoluble carbohydrates. Our goal was to evaluate the effects of replacing the grain portion of the broiler chicken diet with native and extruded wheat and corn bran on growth performance, digestibility, and gut microbiome composition. The research was carried out in 2023 in the vivarium of the Federal Research Centre of Biological Systems and Agrotechnologies of the Russian Academy of Sciences. The Arbor Acres cross broiler chickens (Gallus gallus L.) were divided into four experimental groups (n = 5 each) using the analogue pair method. The birds were fed with complete feed in accordance with the recommendations of the All-Russian Scientific Research and Technological Institute of Poultry Farming (RSRTIPF). Starting from day 21, 10 % of wheat grain (100 g/kg of feed) in the diet of chickens in group I was replaced with wheat bran (B), in group II with bran extrudate (BE), in group III with corn (C), in group IV with corn extrudate (CE). The crude fiber content in the studied components was as follows: 7.7 % for B, 2.7 % for BT, 4.29 % for C, and 1.67 % for CE. Live weight gain was measured by weekly weighing during 21-42 days of life. The chemical composition of diets and droppings was studied. The dry matter content was measured according to GOST 31640-2012 (M., 2012), the mass fraction of crude fat according to GOST 13496.15-2016 (M., 2016), crude protein according to GOST 13496.4-93 (M., 2011), crude fiber according to GOST 31675-2012 (M., 2014). At the end of the experiment, on day 42, the birds were slaughtered. The digestive organs (muscular and glandular stomachs, intestines) were removed, freed from contents and weighed. The contents of the cecum were collected at slaughter. Preparation of DNA libraries, DNA sequencing and bioinformatic processing were performed. Visualization of the bioinformatics data and statistical analysis were carried out using MicrobiomeAnalyst (https://www.microbiomean-alyst.ca/). The resulting OTUs, after filtering and assigning taxonomic affiliation, were used to calculate alpha diversity (Chao1 index, Fisher index, Shannon diversity index, ACE abundance-based coverage estimate, Simpson diversity index; ANOVA statistical method) and beta diversity (NMDS ordination method, distance method, Bray-Curtis index; statistical method PERMANOVA). On day 14 after the start of the experiment, there was a tendency to increase live weight in the groups of chickens receiving extruded products; they had a weight superiority of 8.5 % for EB and 9.9 % for EC compared to similar feeds without extrusion. By the end of the test period, the difference for the same groups in live weight was 8.3 and 10.4 %, respectively, without significance of the differences. The addition of extruded corn to diets led to a statistically significant increase in the digestibility of crude fat (CF): this indicator was 5.1 % (p ≤ 0.05) higher compared to EB and 6.4% (p ≤ 0.05) higher compared to C. The addition of extruded bran to the diet o reduced the digestibility of CF by 3.5 % when compared to group I. When assessing the digestibility of crude protein (CP), the effect was opposite with a statistically significant decrease in groups III and IV, respectively, by 6.6 % (p ≤ 0.05) and 13.2 % (p ≤ 0.05) compared to group I. The remaining indicators of nutrient absorption did not have significant differences. The development of the digestive organs was influenced by the higher fiber content in the native components of bran and corn. The weight of the muscular stomach in group I was 7.02 % (p ≤ 0.001) higher than in birds fed EB, in group III it was 22.04 % (p ≤ 0.05) higher than when fed EC. Intestinal weight decreased in group IV by 22.12 % (p function show_eabstract() { $('#eabstract1').hide(); $('#eabstract2').show(); $('#eabstract_expand').hide(); }
Текст научной статьи Продуктивные качества, переваримость кормов и кишечный микробиом у цыплят-бройлеров (Gallus gallus L.) при добавлении в рацион нативных и экструдированных углеводных компонентов
За 1990-2020 годы в структуре посевов зерновых культур в Российской Федерации произошли существенные изменения, возросла доля продовольственной группы зерновых и снизилась доля фуражной. Затраты на корма составляют до 70 % стоимости производства на птицефабриках, поэтому стоит задача заменить часть пшеницы в рационе более дешевыми компонентами (1) . Для эффективного решения все большее внимание уделяется разработке технологий подготовки кормов для более эффективного использования нутриентов бройлерами (2). Еще 10 лет назад некрахмалистые полисахариды считались антипитательным фактором (3), но положительное действие структурных компонентов клетчатки на пищеварительную систему птицы привлекает все больше внимания (4).
Один из основных факторов, ограничивающих кормовую ценность отрубей для птицы, — повышенное содержание некрахмальных полисахаридов (НКП). В то же время по качеству белка, а именно по аминокислотному составу, пшеничные отруби превосходят зерно пшеницы, а кукуруза имеет богатый углеводный состав (5, 6).
Известно, что экструзия как метод термической и механической обработки, сочетающий высокие температуры и давление, повышает пищевую ценность кормовых ингредиентов и широко используется для улучшения функциональных свойств пищевых продуктов (7). Экструзионное воздействие связано в первую очередь с модификацией трудногидролизуемых углеводов и крахмала в сырье. Изменение крахмала во время высокотемпературной экструзии (от 60 до 140 ° С) происходит на нескольких структурных уровнях, включая распад амилопектина и преобразование кристаллической решетки (8), что влияет на его усвояемость (9). В продуктах с высоким содержанием клетчатки после барогидротермической обработки происходит разрушение полисахаридов, высвобождение фенольных соединений из матрицы и модификация технофункциональных и антиоксидантных свойств (10, 11).
Использование рационов, состоящих из экструзионных компонентов, повышает доступность сложных углеводов для микробиоты, что через действие специфических факторов, регулирующих аппетит (короткоцепочечные жирные кислоты, глюкагоноподобный пептид-1 и пептид YY) предположительно изменяет потребление корма (12). Однако неясно, как рационы, включающие экструдаты, влияют на микробиоту кишечника и физиологию питания цыплят-бройлеров.
Ранее мы подобрали режимы экструзии и оценили изменение углеводного, амино- и жирнокислотного состава отрубей и кукурузы до и после воздействия.
В настоящей работе впервые установлено, что замена до 10 % пшеницы на отруби и кукурузу не снижает продуктивность птицы. Выявлено уменьшение усвоения жира при экструзии отрубей и увеличение этого показателя при обработке кукурузы. Показано, что при разных соотношениях растворимых и нерастворимых углеводов состав микробиом слепой кишки неодинаков.
Наша цель заключалась в оценке влияния замены зерновой части рациона цыплят-бройлеров на нативные и экструдированные пшеничные и кукурузные отруби на показатели роста, переваримость кормов и состав кишечного микробиома.
Методика. Обслуживание животных и экспериментальные исследования выполняли в соответствии с инструкциями и рекомендациями нормативных актов: Модельного закона Межпарламентской Ассамблеи государств-участников Содружества Независимых Государств «Об обращении с животными», статья 20 (постановление МА государств-участников СНГ ¹ 29-17 от 31 октября 2007 года).
Эксперименты проводили в 2023 году в виварии ФНЦ биологических систем и агротехнологий РАН. Цыплят-бройлеров ( Gallus gallus L.) кросса Arbor Acres распределили в четыре опытные группы (в каждой n = 5) методом пар-аналогов.
Птицу кормили полнорационными комбикормами согласно рекомендациям Всероссийского научно-исследовательского и технологического института птицеводства (ВНИТИП). Начиная с 21-х сут, 10 % (100 г/кг рациона) зерна пшеницы в рационе цыплят в I группе заменяли на пшеничные отруби (О), во II группе — на экструдат отрубей (ЭО), в III группе — на кукурузу (К), в IV группе — на экструдат кукурузы (ЭК). Содержание сырой клетчатки в исследуемых компонентах было следующим: О — 7,7 %, ЭО — 2,7 %, К — 4,29 %, ЭК — 1,67 %.
Прирост живой массы учитывали по результатам еженедельного взвешивания (21-42-е сут жизни). Балансовый опыт по оценке переваримости питательных веществ рациона проводили по общепринятой методике (13).
Образцы дробленой кукурузы и пшеничных отрубей предварительно обрабатывали водой до влажности 30 %, перемешивали и отволаживали в течении 24 ч. Обработку проводили на одношнековом прессе-экструдере ПЭШ-30/4 (Россия), максимальная температура головки шнека — в диапазоне +110^+140 ° С, давление — 120 МПа. Полученные образцы сушили на открытом воздухе, размалывали и хранили в пластиковых мешках при температуре не более +8 ° С.
Анализ химического состава рационов и помета проводили в Испытательном центре ЦКП ФНЦ БСТ РАН в трех повторностях. Количество сухого вещества определяли по ГОСТ 31640-2012 (M., 2012), массовую долю сырого жира — по ГОСТ 13496.15-2016 (M., 2016) с использованием автоматического экстрактора жира SER 148/6 («VELP Scientifica», Италия), сырого протеина — по ГОСТ 13496.4-93 (M., 2011) с использованием дигестора DK6 («VELP Scientifica», Италия, полуавтоматическая система дистилляции UDK 139), сырой клетчатки — по ГОСТ 316752012 (M., 2014) с использованием автоматического анализатора клетчатки FIWE Advance («VELP Scientifica», Италия).
Образцы содержимого слепой кишки ( n = 3 от каждой группы) отбирали при убое. Подготовку библиотек ДНК, секвенирование и биоин-форматическую обработку проводили в Центре коллективного научного оборудования «Персистенция микроорганизмов» Института клеточного и внутриклеточного симбиоза УрО РАН (Оренбург), согласно методике (14).
Тотальную ДНК из образцов выделяли комбинированным методом, включавшим механическую гомогенизацию в анализаторе LT («QIAGEN N.V.», Германия) с лизирующей матрицей Y («MP Biomedicals», США). ДНК получали с использованием набора QIAamp Fast DNA Stale Mini Kit («QIAGEN N.V.», Германия) в соответствии с инструкциями производителя. Для исключения возможной контаминации на этапе пробоподготовки использовали отрицательный контроль. Чистоту ДНК контролировали с помощью электрофореза в 1 % агарозном геле и фотометрии на приборе NanoDrop 8000 («ThermoFisher Scientific Inc.», США). Концентрацию ДНК определяли с помощью флуориметра Qubit 4 («Life Technologies», США) с набором для высокочувствительного анализа dsDNA («Life Technologies», США). ДНК-библиотеки для секвенирования были созданы по протоколу Illumina с праймерами к вариабельному участку V3-V4 гена 16S рРНК S-D-Bact-0341-b-S-17 и S-D-Bact-0785-a-A-21 в соответствии с протоколом IlluminaPart #15044223 Rev. B. Библиотеки ДНК были очищены с использованием гранул Agencourt AMPure XP («Beckman Coulter», США) и проверены методом капиллярного электрофореза в усовершенствованной системе Qiaxcel («QIAGEN N.V.», Германия) с набором для скрининга ДНК QIAxcel («QIAGEN N.V.», Германия). Секвенирование проводили на платформе MiSeq («Illumina, Inc.», США) с набором реактивов MiSeqReagent Kit V3 2½300.
Визуализацию результатов биоинформатической обработки и статистический анализ осуществляли с помощью MicrobiomeAnalyst (15). Полученные OTU после фильтрации и определения таксономической принадлежности использовали для расчета показателей альфа-разнообразия (индекс Chao1, индекс Фишера, индекс разнообразия Шеннона, оценка охвата на основе численности ACE, индекс разнообразия Симпсона; статистический метод ANOVA) и бета-разнообразия (метод ординации NMDS, дистанционный метод, индекс Брея-Кертиса; статистический метод PERMANOVA).
Статистический анализ проводили с помощью программ Microsoft Excel («Microsoft», США), Statistica 10.0 («StatSoft, Inc.», США). В статье представлены средние значения показателей ( M ) и стандартные ошибки средних (±SEM). Достоверными считали результаты при р < 0,05.
Результаты. На 14-е сут после начала эксперимента цыплята, получавшие экструдированные продукты, имели превосходство по массе на 8,5 % для ЭО и 9,9 % для ЭК в сравнении с получавшими аналогичные корма без экструзии. К концу учетного периода разница для тех же групп по живой массе сохранялась и составила соответственно 8,3 и 10,4 % (табл. 1).
1. Живая масса (г) цыплят-бройлеров ( Gallus gallus L.) кросса Arbor Acres при использовании в рационе нативных и экструдированных компонентов (по n = 5, M ±SEM; опыт в условиях вивария ФНЦ биологических систем и агротехнологий РАН, г. Оренбург, 2023 год)
|
Период |
Группа |
|||
|
I (О) |
п II (ЭО) 1 |
III (К) |
IV (ЭК) |
|
|
Начало опыта |
663,2±20,3 |
664,0±24,2 |
664,2±30,8 |
663,4±30,1 |
|
1-я нед |
1041,2±21,7 |
1062,8±58,7 |
1101,0±31,2 |
1135,0±44,7 |
|
2-я нед |
1582,6±49,3 |
1717,5±43,6 |
1610,2±55,9 |
1770,8±63,5 |
|
3-я нед |
2238,6±42,6 |
2412,5±82,6 |
2107,8±178,4 |
2439,3±44,1 |
|
Окончание опыта |
2391,6±38,1 |
2590,0±98,0 |
2298,5±50,8 |
2537,0±75,3 |
Примечание. О — отруби, ЭО — экструдат отрубей, К — кукуруза, ЭК — экструдат кукурузы.
Повышенная относительно других групп интенсивность роста подопытной птицы, получавшей в составе рациона экструдаты, обусловила их превосходство по абсолютному приросту живой массы за период эксперимента (табл. 2). По этому показателю цыплята-бройлеры во II группе опережали I группу на 11,4 %, а в IV группе абсолютный прирост был выше на 14,6 % относительно III группы. Общий расход кормов на единицу прироста живой массы имел наименьшее значение в группе К — 1,68 кг.
2. Показатели продуктивности и поедаемость корма цыплятами-бройлерами ( Gallus gallus L.) кросса Arbor Acres при использовании в рационе нативных и экструдированных компонентов (по n = 5, M ±SEM; опыт в условиях вивария ФНЦ биологических систем и агротехнологий РАН, г. Оренбург, 2023 год)
|
Показатель |
Группа |
|||
|
I (О) |
II (ЭО) 1 |
III (К) 1 |
IV (ЭК) |
|
|
Прирост живой массы, г |
1728,4±38,1 |
1926,0±97,9 |
1634,3±50,8 |
1873,6±75,1 |
|
Поедаемость корма за эксперимент, г/гол |
3177,8 |
3785,4 |
2752,6 |
3568,9 |
|
Затраты корма на 1 кг прироста живой массы, кг |
1,84 |
1,97 |
1,68 |
1,90 |
Примечание. О — отруби, ЭО — экструдат отрубей, К — кукуруза, ЭК — экструдат кукурузы.
Замена зерновой части рациона в количестве 100 г/кг нативными и экструдированными отрубями и кукурузой не привела к статистически значимым различиям по показателям роста и развития цыплят-бройлеров, что согласуется с ранее описанными результатами (16). Поедаемость кормов была лучшей в группах с экструдированными компонентами, поскольку экструзия улучшает вкусовые качества сырья и снижает содержание непе-ревариваемой клетчатки (17).
Внесение в рационы экструдированной кукурузы приводило к статистически значимому увеличению переваримости сырого жира (СЖ): этот показатель был больше на 5,1 % (р ≤ 0,05), чем в группе с ЭО, и на 6,4 % (р ≤ 0,05) — при сравнении с К. Внесение в рацион цыплят-бройлеров экструдированных отрубей снижало переваримость СЖ на 3,5 % по сравнению с I группой (табл. 3).
3. Коэффициенты переваримости питательных веществ рациона (%) цыплят-бройлеров ( Gallus gallus L.) кросса Arbor Acres при использовании нативных и экструдированных компонентов (по n = 5, M ±SEM; опыт в условиях вивария ФНЦ биологических систем и агротехнологий РАН, г. Оренбург, 2023 год)
а, b, с Различия при сравнении опытных групп соответственно с I, II и III группами статистически значимы при р ≤ 0,05.
При оценке переваримости сырого протеина (СП) отмечали обратный эффект: статистически значимое снижение в III и IV группах соответственно на 6,6 % (р ≤ 0,05) и 13,2 % (р ≤ 0,05) по сравнению с I группой. При сравнении с III группой, получавшей нативную кукурузу, переваримость СП в IV группе была ниже на 6,6 %. Во II группе, которой скармливали ЭО, переваримость СП была на 8,2 % выше, чем при добавлении в рацион ЭК.
Остальные показатели усвоения питательных веществ не имели достоверно значимых различий. Была отмечена тенденция к снижению переваримости сухого вещества во всех опытных группах по сравнению с I группой, получавшей О. Так, при скармливании ЭО переваримость СВ рациона у цыплят составляла 72,06 %, что было на 4,7 % ниже при сравнении с I группой. Важно отметить, что при включении в рацион ЭО и ЭК переваримость СВ оказалась одинаковой.
Данные по усвояемости питательных веществ в нашем исследовании были противоречивы, показатели зависели от структуры используемой добавки. Например, включение экструдированных пшеничных отрубей не изменяло усвояемость СЖ, наилучшие показатели наблюдались при введении кукурузы в обработанном виде — увеличение на 6,4 % (р < 0,05) при сравнении с К. Некоторые исследователи утверждают, что процесс экструзии способен повысить переваривание жира (18-20). Напротив, F.G. Boroojeni с соавт. (21) сделали вывод об отсутствии положительной связи между экструзионной обработкой корма и усвояемостью жира и белка.
Усвоение протеина оказалось более эффективным в группах, получавших нативные и экструдированные пшеничные отруби (р < 0,05), чем в группах, получавших кукурузу. Однако в проведенном эксперименте предварительная экструзионная обработка как отрубей, так и кукурузы не повышала усвояемость СП, что отличается от данных A. Ahmed с соавт. (22), H.S. Faridah с соавт. (17) и M. Hejdysz с соавт. (23). В нашем случае компоненты с большим содержанием клетчатки увеличили этот показатель, что можно объяснить влиянием отрубей на развитие желудочно-кишечного тракта и сопутствующим увеличением секреции пищеварительных ферментов (3).
Переваримость клетчатки в нашем эксперименте не зависела от ее количества в исследуемых компонентах, что согласуется с имеющимися данными (24).
Полученные результаты усвоения питательных веществ по группам могут быть связаны со многими факторами, такими как источник клетчатки, состав основного рациона, при этом эффекты становятся более выраженными, когда к основным рационам добавляется умеренное количество нерастворимой клетчатки (25).
4. Масса органов желудочно-кишечного тракта (г/кг живой массы) у цыплят-бройлеров ( Gallus gallus L.) кросса Arbor Acres при использовании в рационе нативных и экструдированных компонентов (по n = 5, M ±SEM; опыт в условиях вивария ФНЦ биологических систем и агротехнологий РАН, г. Оренбург, 2023 год)
b Различия при сравнении опытных групп со II группой статистически значимы при р < 0,0.
с, сс Различия при сравнении опытных групп с III группой статистически значимы соответственно при р < 0,05 и p < 0,01.
Также представляют интерес изменения микробиома содержимого слепой кишки, где происходят процессы преобразования сложных соединений, входящих в состав НКП изучаемых компонентов. В результате секвенирования ДНК из образцов содержимого слепой кишки цыплят из I, II, III и IV групп получили соответственно 35608, 46453, 29316 и 42354 ридов. Число OTUs (operational taxonomic units) по группам составило соответственно 272, 266, 401 и 382. Полученные OTUs были таксономически сгруппированы от уровня филума до уровня рода (табл. 5).
-
5. Число таксонов различного уровня, идентифицированных в микробиоме кишечника цыплят-бройлеров ( Gallus gallus L.) кросса Arbor Acres при использовании в рационе нативных и экструдированных компонентов (по n = 3, M ±SEM; опыт в условиях вивария ФНЦ биологических систем и агротехнологий РАН, г. Оренбург, 2023 год)
|
Таксон |
Группа |
|||
|
I (О) |
II (ЭО) |
III (К) |
IV (ЭК) |
|
|
Филум |
9±2,56 |
8±3,60 |
9±3,30 |
9±2,90 |
|
Класс |
15±5,05 |
14±5,40 |
16±5,98 |
16±4,89 |
|
Отряд |
19±6,60 |
17±5,50 |
20±6,98 |
21±7,05 |
|
Семейство |
31±12,50 |
30±11,30 |
34±12,30 |
34±12,90 |
|
Род |
82±27,60 |
82±29,40 |
93±30,10 |
93±30,50 |
Примечание. О — отруби, ЭО — экструдат отрубей, К — кукуруза, ЭК — экструдат кукурузы.
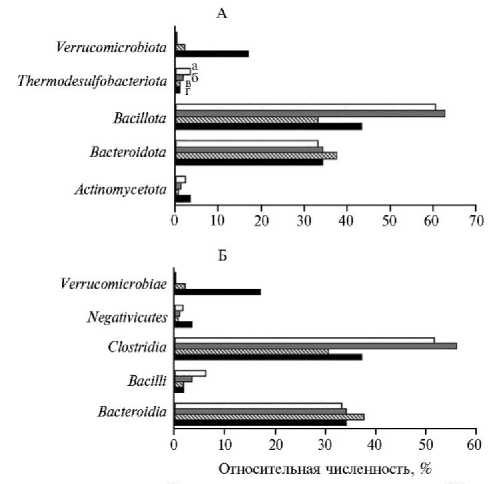
Рис. 1. Основные таксономические группы бактерий на уровне филумов (А) и классов (Б) в микробиоте слепой кишки у цыплят-бройлеров ( Gallus gallus L.) кросса Arbor Acres при использовании в рационе нативных и экструдированных компонентов: а — I группа (скармливание отрубей); б — II группа (скармливание экструдата отрубей); в — III группа (скармливание кукурузы); г — IV группа (скармливание экструдата кукурузы) (по n = 3; опыт в условиях вивария ФНЦ биологических систем и агротехнологий РАН, г. Оренбург, 2023 год) .
При анализе состава микробиоты слепой кишки цыплят-бройлеров была обнаружена взаимосвязь между содержанием некоторых таксонов и используемым рационом. Так, основу микробиома слепой кишки цыплят, получавших пшеничные отруби (рис. 1), составляли Bacteroidota , Bacillota и
Verrucomicrobiota . Микроорганизмы, относящиеся к классам Bacteroidia и Clostridia , составляли более 70 % от общего числа идентифицированных бактерий. Эти классы были представлены в основном семействами Bacteroidaceae , Rikenellaceae , Lachnospiraceae и Oscillospiraceae . Филум Verrucomicrobiota был представлен единственным семейством Akkermansiaceae . Наиболее многочисленными представителями микробиоты на уровне рода были Phocaeicola , Alistipes , Rikenella , unclassified Oscillospiraceae и Akkermansia .
Включение кукурузы в корм цыплят-бройлеров привело к тому, что основными таксономическими группами в микробиоме слепой кишки оказались Bacteroidota и Bacillota. Число микроорганизмов, относящихся к фи-луму Verrucomicrobiota , составило менее 1 % (см. рис. 1).
Таксономический состав микробиоты слепой кишки у цыплят-бройлеров, получавших с кормом ЭК, по численному соотношению основных бактериальных групп был практически аналогичен таковому у птицы, получавшей кукурузу без предварительно экструзии. В то же время ЭК в рационе цыплят способствовала увеличению доли микроорганизмов рода Bacteroides и уменьшению доли бактерий, относящихся к unclassified Oscillospiraceae .
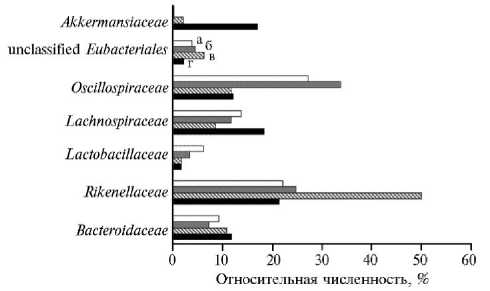
Рис. 2. Основные таксономические группы на уровне семейств бактерий в микробиоте слепой кишки у цыплят-бройлеров ( Gallus gallus L.) кросса Arbor Acres при использовании в рационе нативных и экструдированных компонентов: а — I группа (скармливание отрубей); б — II группа (скармливание экструдата отрубей); в — III группа (скармливание кукурузы); г — IV группа (скармливание экструдата кукурузы) (по n = 3; опыт в условиях вивария ФНЦ биологических систем и агротехнологий РАН, г. Оренбург, 2023 год) .
В отличие от I группы, в III группе микробиом слепой кишки цыплят характеризовался большим числом бактерий семейств Rikenellaceae , Oscil-lospiraceae и Lactobacillaceae (рис. 2). Напротив, в слепой кишке цыплят из II группы отмечалась меньшая представленность бактерий семейств Bac-teroidaceae и Lachnospiraceae .
При добавлении экструдированных пшеничных отрубей в рацион число бактерий филумов Verrucomicrobiota и Bacillota снижалось, микроорганизмов таксона Bacteroidota — повышалось. Число бактерий семейства Rikenellaceae стало на 28 % больше, чем при применении отрубей без предварительной экструзии, вследствие большей представленности микроорганизмов родов Alistipes и Rikenella .
Анализ микробиоты на уровне рода показал, что 42,2 % бактерий относились к Alistipes и unclassified Oscillospiraceae (рис. 3).
Сравнение индексов альфа-разнообразия микробиоты кишечника цыплят-бройлеров из I и II групп, а также из III и IV групп не выявило значительных различий (табл. 6). В то же время необходимо отметить, что при включении кукурузы (как с экструзией, так и без) в рацион индексы Chao1 были выше, чем при использовании пшеничных отрубей
(как с экструзией, так и без), что может свидетельствовать о большем числе таксонов на уровне рода.
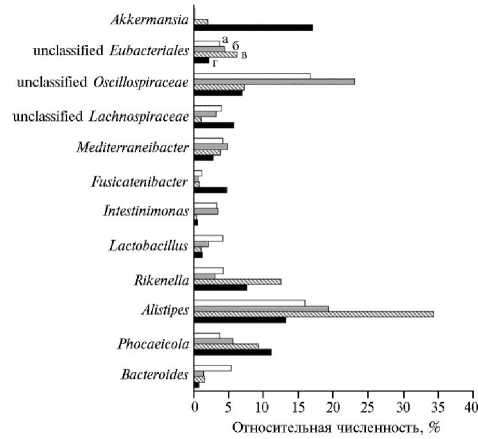
Рис. 3. Основные таксономические группы на уровне родов бактерий в микробиоте слепой кишки у цыплят-бройлеров ( Gallus gallus L.) кросса Arbor Acres при использовании в рационе нативных и экструдированных компонентов: а — I группа (скармливание отрубей); б — II группа (скармливание экструдата отрубей); в — III группа (скармливание кукурузы); г — IV группа (скармливание экструдата кукурузы) ( n = 3; опыт в условиях вивария ФНЦ биологических систем и агротехнологий РАН, г. Оренбург, 2023 год) .
-
6. Индексы альфа-разнообразия микробиоты слепой кишки цыплят-бройлеров ( Gallus gallus L.) кросса Arbor Acres при использовании в рационе нативных и экструдированных компонентов ( n = 3, M ±SEM; опыт в условиях вивария ФНЦ биологических систем и агротехнологий РАН, г. Оренбург, 2023 год)
|
Группа Показатель II (ЭО) руппаIII (К) IV (ЭК) |
p-value |
|
Индекс Chao1 216,5±13,50 212,5±2,50 317±11 297,5±4,50 ACE 223,7±13,60 219,7±2,49 325,6±10,90 306,0±4,54 Индекс Фишера 33,8±1,05 32,4±0,35 51,2±1,23 48,7±0,14 Индекс разнообразия Симпсона 0,89±0,030 0,85±0,060 0,97±0,001 0,97±0,005 Индекс разнообразия Шеннона 3,4±0,12 3,4±0,49 4,4±0,06 4,3±0,07 |
0,03 0,19 0,16 0,28 0,13 |
Примечание. О — отруби, ЭО — экструдат отрубей, К — кукуруза, ЭК — экструдат кукурузы; ACE — оценка охвата на основе численности.
Оценка бета-разнообразия выявила значимые различия между микробиомами слепой кишки у цыплят-бройлеров из I и II групп (рис. 4).
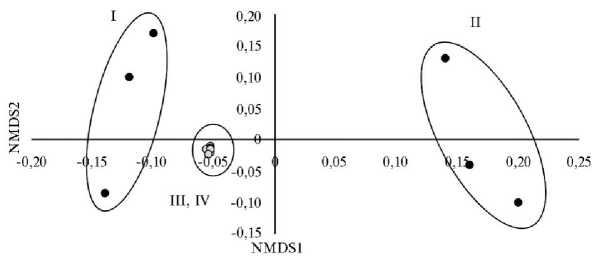
Рис. 4. Бета-разнообразие микробиты слепого отростка кишечника у цыплят-бройлеров (Gallus gallus L.) кросса Arbor Acres при использовании в рационе нативных и экструдированных компонентов (статистический метод PERMANOVA, неметрическое многомерное масштабирование, несходство Брея-Кертис): а — I группа (скармливание отрубей); б — II группа (скармливание экструдата отрубей); в — III группа (скармливание кукурузы); г — IV группа (скармливание экструдата кукурузы) (по n = 3; опыт в условиях вивария ФНЦ биологических систем и агротехнологий РАН, г. Оренбург, 2023 год).
Также были показаны различия между микробиомами цыплят, получавших кукурузу и пшеничные отруби в любом виде. Напротив, различий у цыплят-бройлеров из III и IV групп выявлено не было.
Из-за отсутствия некоторых ферментов процессы в пищеварительной системе птицы напрямую зависят от функционирования микробиома слепой кишки (28). Именно в этом отделе происходит расщепление целлюлозы и крахмала. Состав микрофлоры слепой кишки цыплят-бройлеров при применении кормовых добавок c различной структурой НКП позволяет судить об их эффективности (29).
Использование компонентов углеводной природы, имеющих разные физико-химические свойства, в рационе цыплят-бройлеров приводит к формированию схожего таксономического состава микробиома слепой кишки, в котором отмечалось присутствие ряда типичных представителей (30). В то же время в микробиоме цыплят-бройлеров, получавших кукурузу или пшеничные отруби, были выявлены особенности, связанные с количественным соотношением некоторых таксонов. Аналогичные результаты представлены и в других сообщениях (31).
Особенностью микробиома слепой кишки цыплят-бройлеров, получавших в рационе пшеничные отруби без предварительной экструзии, было присутствие бактерий рода Akkermansia (16 % от общего числа идентифицированных микроорганизмов). Есть данные о высокой корреляции между потреблением птицей пшеничных отрубей и повышенной численностью бактерий Akkermansia (32). Некоторым представителям рода Akkermansia отводится важная роль в поддержании нормального состояния слизистой оболочки кишечника, кроме того, их рассматривают в качестве потенциальных пробиотических микроорганизмов (33, 34). В остальных опытных группах доля бактерий этого рода не превышала 2 % от общего числа. Снижение численности бактерий Akkermansia в микробиоме кишечника цыплят-бройлеров, получавших ЭО, вероятно, обусловлено изменениями химического состава пшеничных отрубей в результате экструзии.
Сообщалось о связи между численностью Akkermansia в кишечнике и высоким содержанием клетчатки в корме (35). В нашей работе экструзия приводила к снижению массовой доли сырой клетчатки (5). В ряде исследований на цыплятах-бройлерах была показана взаимосвязь между увеличением численности Akkermansia в микробиоте кишечника и улучшением некоторых физиологических параметров (36, 37). В то же время обнаружена отрицательная корреляция между долей бактерий рода Akkermansia и живой массой цыплят-бройлеров (38). В нашем исследовании меньшая численность Akkermansia в микробиоме слепой кишки отмечалась в группах цыплят с более высокими показателями прироста живой массы (II и IV группы).
Примечательно, что при использовании ЭО увеличивалась доля бактерий семейства Rikenellaceae (род Alistipes ) в сравнении с группой, получавшей нативные отруби. В слепой кишке цыплят бактерии Alistipes осуществляют синтез короткоцепочечных жирных кислот (ацетата, пропионата, сукцината), которые оказывают противовоспалительное действие и участвуют в синтезе метаболитов липидного обмена в печени (39). Сукцинат — основной продукт ферментации бактерий Alistipes и важный источник энергии в кишечнике, что коррелирует с увеличением живой массы цыплят.
В исследовании C. De Maesschalck с соавт. (40) отмечается, что более высокому относительному содержанию Alistipes в микробиоме слепой кишки бройлеров способствует использование более доступной формы целлюлозы (аморфная целлюлоза). Наличие подобных взаимосвязей дает возможность предположить, что экструзия позволяет сделать компоненты корма более доступными для ферментации и облегчает процесс пищеварения.
По сравнению с рационами на основе пшеничных отрубей, корма с кукурузой повышали численность бактерий семейства Oscillospiraceae (наиболее многочисленная группа unclassified Oscillospiraceae ). Микроорганизмы семейства Oscillospiraceae , как и рода Alistipes , — целлюлотические бактерии, и увеличение их доли в составе микробиоты при использовании в рационе кукурузы, вероятно, обусловлено более высокой массовой долей кукурузного крахмала (28).
Список литературы Продуктивные качества, переваримость кормов и кишечный микробиом у цыплят-бройлеров (Gallus gallus L.) при добавлении в рацион нативных и экструдированных углеводных компонентов
- Масалов В.Н., Березина Н.А., Червонова И.В. Состояние зернового хозяйства России, роль зерновых в кормлении сельскохозяйственных животных и питании человека. Вестник аграрной науки, 2021, 2(89): 3-15 (doi: 10.17238/issn2587-666X.2021.2.3).
- Sozcu A. Growth performance, pH value of gizzard, hepatic enzyme activity, immunologic indi-cators, intestinal histomorphology, and cecal microflora of broilers fed diets supplemented with processed lignocellulose. Poultry Science, 2019, 98(12): 6880-6887 (doi: 10.3382/ps/pez449).
- Mateos G.G., Jiménez-Moreno E., Serrano M.P., Lázaro R.P. Poultry response to high levels of dietary fiber sources varying in physical and chemical characteristics. Journal of Applied Poultry Research, 2012, 21(1): 156-174 (doi: 10.3382/japr.2011-00477).
- Jha R., Fouhse J.M., Tiwari U.P., Li L., Willing B.P. Dietary fiber and intestinal health of monogastric animals. Front. Vet. Sci., 2019, 6: 48 (doi: 10.3389/fvets.2019.00048).
- Холодилина Т.Н., Климова Т.А., Курилкина М.Я., Ваншин В.В. Влияние баротермиче-ской обработки компонентов рациона на микробное сообщество кишечника цыплят-бройлеров. Животноводство и кормопроизводство, 2022, 105(1): 92-102 (doi: 10.33284/2658-3135-105-1-92).
- Холодилина Т.Н., Курилкина М.Я., Атландерова К.Н. Экструзионная обработка как фактор, определяющий аминокислотный состав различных компонентов корма для цыплят-бройлеров. Животноводство и кормопроизводство, 2022, 105(1): 74-81 (doi: 10.33284/2658-3135-105-1-74).
- Rojas O.J., Stein H.H. Processing of ingredients and diets and effects on nutritional value for pigs. J. Anim. Sci. Biotechnol., 2017, 8: 48 (doi: 10.1186/s40104-017-0177-1).
- Li M., Hasjim J , Xie F., Halley P.J., Gilbert R.G. Shear degradation of molecular, crystalline, and granular structures of starch during extrusion. Starch - Stärke, 2014, 66(7-8): 595-605 (doi: 10.1002/star.201300201).
- Zhang Z., Zhu M., Xing B., Liang Y., Zou L., Li M., Fan X., Ren G., Zhang L., Qin P. Effects of extrusion on structural properties, physicochemical properties and in vitro starch digestibility of Tartary buckwheat flour. Food Hydrocolloids, 2023, 135(10): 108197 (doi: 10.1016/j.foodhyd.2022.108197).
- Garcia-Amezquita L.E., Tejada-Ortigoza V., Pérez-Carrillo E., Serna-Saldívar S.O., Campan-ella O.H., Welti-Chanes J. Functional and compositional changes of orange peel fiber thermally-treated in a twin extruder. LWT, 2019, 111: 673-681 (doi: 10.1016/j.lwt.2019.05.082).
- Villasante J., Pérez‐Carrillo E., Heredia‐Olea E., Metón I., Almajano M.P. In vitro antioxidant activity optimization of nut shell (Carya illinoinensis) by extrusion using response surface methods. Biomolecules, 2019, 9(12): 883 (doi: 10.3390/biom9120883).
- Ringseis R., Gessner D.K., Eder K. The gut-liver axis in the control of energy metabolism and food intake in animals. Annual Review of Animal Biosciences, 2020, 8: 295-319 (doi: 10.1146/an-nurev-animal-021419-083852).
- Егоров И.А., Манукян В.А., Ленкова Т.Н., Околелова Т.М., Лукашенко В.С., Шевяков А.Н., Игнатова Г.В., Егорова Т.В., Андрианова Е.Н., Розанов Б.Л., Лысенко М.А., Егорова Т.А., Грозина А.А., Лаптев Г.Ю., Никонов И.Н., Александрова И.Л., Ильина Л.А., Новикова Н.И. Методика проведения научных и производственных исследований по кормлению сельскохозяйственной птицы. Молекулярно-генетические методы определения микрофлоры кишечника. Сергиев Посад, 2013.
- Sizova E., Yausheva E., Marshinskaia O., Kazakova T., Khlopko Y., Lebedev S. Elemental com-position of the hair and milk of black-spotted cows and its relationship with intestinal microbiome reorganization. Veterinary World, 2022, 15(11): 2565-2574 (doi: 10.14202/vetworld.2022.2565-2574).
- Dhariwal A., Chong J., Habib S., King I.L., Agellon L.B., Xia J. MicrobiomeAnalyst: a web-based tool for comprehensive statistical, visual and meta-analysis of microbiome data. Nucleic Acids Research, 2017, 45(W1): W180-W188 (doi: 10.1093/nar/gkx295).
- Adewole D. Effect of dietary supplementation with coarse or extruded oat hulls on growth per-formance, blood biochemical parameters, ceca microbiota and short chain fatty acids in broiler chickens. Animals, 2020, 10(8): 1429 (doi: 10.3390/ani10081429).
- Faridah H.S., Goh Y.M., Noordin M.M., Liang J.B. Extrusion enhances apparent metabolizable energy, ileal protein and amino acid digestibility of palm kernel cake in broilers. Asian-Australasian Journal of Animal Sciences, 2020, 33(12): 1965-1974 (doi: 10.5713/ajas.19.0964).
- Hejdysz M., Kaczmarek S.A., Kubiś M., Adamski M., Perz K., Rutkowski A. The effect of faba bean extrusion on the growth performance, nutrient utilization, metabolizable energy, excretion of sialic acids and meat quality of broiler chickens. Animal, 2019, 13(8): 1583-1590 (doi: 10.1017/S175173111800366X).
- Oryschak M., Korver D., Zuidhof M., Meng X., Beltranena E. Comparative feeding value of extruded and nonextruded wheat and corn distillers dried grains with solubles for broilers. Poultry Science, 2010, 89(10): 2183-2196 (doi: 10.3382/ps.2010-00758).
- Rutkowski A., Kaczmarek S.A., Hejdysz M., Jamroz D. Effect of extrusion on nutrients digesti-bility, metabolizable energy and nutritional value of yellow lupine seeds for broiler chickens. Annals of Animal Science, 2016, 16(4): 1059-1072 (doi: 10.1515/aoas-2016-0025).
- Boroojeni F.G., Svihus B., von Reichenbach H.G., Zentek J. The effects of hydrothermal processing on feed hygiene, nutrient availability, intestinal microbiota and morphology in poultry — a review. Animal Feed Science and Technology, 2016, 220: 187-215 (doi: 10.1016/j.anifeedsci.2016.07.010).
- Ahmed A., Zulkifli I., Farjam A.S., Abdullah N., Liang J.B. Extrusion enhances metabolizable energy and ileal amino acids digestibility of canola meal for broiler chickens. Italian Journal of Animal Science, 2014, 13(1): 3032 (doi: 10.4081/ijas.2014.3032).
- Hejdysz M., Kaczmarek S.A., Adamski M., Rutkowski A. Influence of graded inclusion of raw and extruded pea (Pisum sativum L.) meal on the performance and nutrient digestibility of broiler chick-ens. Animal Feed Science and Technology, 2017, 230: 114-125 (doi: 10.1016/j.anifeedsci.2017.05.016).
- Shang Q., Wu D., Liu H., Mahfuz S., Piao X. The impact of wheat bran on the morphology and physiology of the gastrointestinal tract in broiler chickens. Animals, 2020, 10(10): 1831 (doi: 10.3390/ani10101831).
- Jiménez-Moreno E., de Coca-Sinova A., González-Alvarado J.M., Mateos G.G. Inclusion of insoluble fiber sources in mash or pellet diets for young broilers. 1. Effects on growth performance and water intake. Poultry Science, 2016, 95(1): 41-52 (doi: 10.3382/ps/pev309).
- Jiménez-Moreno E., González-Alvarado J.M., de Coca-Sinova A., Lázaro R.P., Cámara L., Mateos G.G. Insoluble fiber sources in mash or pellets diets for young broilers. 2. Effects on gastrointestinal tract development and nutrient digestibility. Poultry Science, 2019, 98(6): 2531-2547 (doi: 10.3382/ps/pey599).
- Guzmán P., Saldaña B., Kimiaeitalab M.V., García J., Mateos G.G. Inclusion of fiber in diets for brown-egg laying pullets: effects on growth performance and digestive tract traits from hatching to 17 weeks of age. Poultry Science, 2015, 94(11): 2722-2733 (doi: 10.3382/ps/pev288).
- Conlon M.A., Bird A.R. The impact of diet and lifestyle on gut microbiota and human health. Nutrients, 2015, 7(1): 17-44 (doi: 10.3390/nu7010017).
- Фисинин В.И., Ильина Л.А., Йылдырым Е.А., Никонов И.Н., Филиппова В.А., Лап-тев Г.Ю., Новикова Н.И., Грозина А.А., Ленкова Т.Н., Манукян В.А., Егоров И.А. Бакте-риальное сообщество слепых отростков кишечника цыплят-бройлеров на фоне питатель-ных рационов различной структуры. Микробиология, 2016, 85(4): 472-480.
- Xiao Y., Xiang Y., Zhou W., Chen J., Li K., Yang H. Microbial community mapping in intestinal tract of broiler chicken. Poultry Science, 2017, 96(5): 1387-1393 (doi: 10.3382/ps/pew372).
- Borda-Molina D., Mátis G., Mackei M., Neogrády Z., Huber K., Seifert J., Camarinha-Silva A. Caeca microbial variation in broiler chickens as a result of dietary combinations using two cereal types, supplementation of crude protein and sodium butyrate. Front. Microbiol., 2021, 11: 617800 (doi: 10.3389/fmicb.2020.617800).
- Molnár A., Such N., Farkas V., Pál L., Menyhárt L., Wágner L., Husvéth F., Dublecz K. Effects of wheat bran and Clostridium butyricum supplementation on cecal microbiota, short-chain fatty acid concentration, pH and histomorphometry in broiler chickens. Animals, 2020, 10(12): 2230 (doi: 10.3390/ani10122230).
- Zhou K. Strategies to promote abundance of Akkermansia muciniphila, an emerging probiotics in the gut, evidence from dietary intervention studies. Journal of Functional Foods, 2017, 33: 194-201 (doi: 10.1016/j.jff.2017.03.045).
- Hagi T., Belzer C. The interaction of Akkermansia muciniphila with host-derived substances, bac-teria and diets. Appl Microbiol Biotechnol., 2021, 105(12): 4833-4841 (doi: 10.1007/s00253-021-11362-3).
- Yao S., Zhao Y., Chen H., Sun R., Chen L., Huang J., Yu Z., Chen S. Exploring the plasticity of diet on gut microbiota and its correlation with gut health. Nutrients, 2023, 15(15): 3460 (doi: 10.3390/nu15153460).
- Yan W., Sun C., Yuan J., Yang N. Gut metagenomic analysis reveals prominent roles of Lacto-bacillus and cecal microbiota in chicken feed efficiency. Sci. Rep., 2017, 7: 45308 (doi: 10.1038/srep45308).
- Cui X., Gou Z., Jiang Z., Li L., Lin X., Fan Q., Wang Y., Jiang S. Dietary fiber modulates abdominal fat deposition associated with cecal microbiota and metabolites in yellow chickens. Poultry Science, 2022, 101(4): 101721 (doi: 10.1016/j.psj.2022.101721).
- Han G.G., Kim E.B., Lee J., Lee J.-Y., Jin G., Park J., Huh C.-S., Kwon I.K., Kil D.Y., Choi Y.J., Kong C. Relationship between the microbiota in different sections of the gastrointes-tinal tract, and the body weight of broiler chickens. SpringerPlus, 2016, 5(1): 911 (doi: 10.1186/s40064-016-2604-8).
- Zhang B., Liu N., Hao M., Zhou J., Xie Y., He Z. Plant-derived polysaccharides regulated immune status, gut health and microbiota of broilers: a review. Front. Vet Sci., 2022, 8: 791371 (doi: 10.3389/fvets.2021.791371).
- De Maesschalck C., Eeckhaut V., Maertens L., De Lange L., Marchal L., Daube G., Dewulf J., Haesebrouck F., Ducatelle R., Taminau B., Van Immerseel F. Amorphous cellulose feed supple-ment alters the broiler caecal microbiome. Poultry Science, 2019, 98(9): 3811-3817 (doi: 10.3382/ps/pez090).
