Различия в механизмах перехода к земледелию в лесной зоне Европы и Северной Америки

Автор: Леммен К.
Журнал: Археология, этнография и антропология Евразии @journal-aeae-ru
Рубрика: Палеоэкология. Каменный век
Статья в выпуске: 3 (55), 2013 года.
Бесплатный доступ
Несмотря на сходные природные условия лесной зоны, возникновение и распространение производящего хозяйства в восточной части Северной Америки и Европе происходили по-разному. Для объяснения различий в сроках, скорости и механизмах перехода в работе используется моделирующая система мирового землепользования и технологической эволюции -численная модель для моделирования демографии, инноваций, доместикации, миграций и торговли в геоэкологическом контексте. Показано, что европейское население получило комплекс окультуренных видов растений и животных с других территорий и быстро перешло к производящему хозяйству. В отличие от Европы, в восточной части Северной Америки охота-собирательство и земледелие-скотоводство сосуществовали в течение длительного времени, а земледелие медленно интегрировалось в уже существующую систему жизнеобеспечения. Из этого делается вывод о качественном экономическом различии в переходе к земледелию в данных регионах: ограниченных людских ресурсах в Европе и ограниченных природных ресурсах в восточной части Северной Америки.
Раннее земледелие, "неолитический пакет", население доколумбовой америки, адаптивная динамика, социально-технологическая модель, экодинамика человека
Короткий адрес: https://sciup.org/14522999
IDR: 14522999 | УДК: 930.85
Текст научной статьи Различия в механизмах перехода к земледелию в лесной зоне Европы и Северной Америки
Возникновение земледелия в лесной зоне
Первая неолитическая революция произошла в Восточном Средиземноморье и долинах китайских рек Янцзы и Хуанхэ между 10000 и 8000 г. до н.э., когда охота и собирательство сменились на земледелие и скотоводство как основные способы добывания пищи [Willcox, 2005; Kuijt, Goring-Morris, 2002; Londo et al., 2006]. Впоследствии почти каждое человеческое сообщество во всем мире в разное время, с разной скоростью и с помощью различных механизмов переходило к производящему хозяйству. В ряде регионов этот переход не был самостоятельным: методы хозяйствования перенимались из основных земледельческих и скотоводческих центров (см., напр.: [Wen et al., 2004; Fuller, 2011; Lemmen, Gronenborn, Wirtz, 2011]). Примерами могут служить Европа и восточная часть Северной Америки (о последней см.: [Price et al., 2001; Zeder, 2008; Smith, 2011]).
На большей части Европы переход к производящему хозяйству произошел между 6000 и 4000 г. до н.э. с плавным изменением сроков с юго-востока (самое раннее в Греции и на Балканах [Perlès, 2001]) к северо-западу (Южная Скандинавия [Price, 2003] и Британия [Whittle, Cummings, 2007]). Наиболее ярким примером крупного однородного земледельческого комп-
Археология, этнография и антропология Евразии 3 (55) 2013 © Леммен К., 2013
лекса является культура линейно-ленточной керамики Центральной Европы [Lüning, 2005]. Всюду возникновению производящего хозяйства сопутствовало появление керамических изделий. Ни один вид растений не был культивирован в Европе; все возделывавшиеся там культуры родом из Западной Азии, где они впервые появились. Проникший в Европу «неолитический пакет» состоял из окультуренных видов растений и животных, происхождение которых связано с регионом «Плодородного полумесяца» [Flannery, 1973], и включал пшеницу, ячмень, рожь, чечевицу, горох; крупный рогатый скот, овец, коз и свиней [Willcox, 2005; Luikart et al., 2001; Edwards et al., 2007; Larson et al., 2007; Zeder, 2008]. Хотя генетический анализ свидетельствует о миграционных процессах, сопровождавших появление «неолитического пакета» [Haak et al., 2010], исследование моделей культурной экспансии показывает столь же большую вероятность сценария диффузионного распространения этих видов растений и животных в Европе [Ackland et al., 2007; Lemmen, Gronenborn, Wirtz, 2011].
Переход к производящему хозяйству в Северной Америке произошел намного позже, между 1000 г. до н.э. и 1000 г. н.э. Для ее восточной части этот период обозначается как вудлендский. Местные окультуренные растения, такие как амарант и подсолнечник, возделывались параллельно с охотой и собирательством как основными формами хозяйственной деятельности. Постепенно на восток Северной Америки из Мексики проникали кукуруза и бобы, становясь основными культурами; одомашненные животные отсутствовали [Piperno, 2011]. Вудлендский период характеризуется постоянным использованием керамики и постепенным усилением зависимости от земледелия [Anderson, Mainfort, 2002]. В отличие от Европы, культурные растения распространялись в восточной части Северной Америки через торговлю, а не посредством миграции населения из регионов окультуривания; культивацию осваивало местное население [Hart, 1999].
Для успешного выращивания урожая необходимы подходящие климатические условия, а также достаточное количество возделываемых почв и воды. За исключением безводных плоскогорий вдоль Западных Кордильер Северная Америка и Европа имеют сходные климат, типы растительности, топографию и обладают ландшафтным разнообразием, включающим крупные реки, горные хребты, равнины и холмы. Естественный тип растительности на большей части их лесных зон представлен лесами умеренного пояса [Williams et al., 2000; Thompson, Anderson, 2000; Cheddadi et al., 1997].
В данной работе используется модель мирового землепользования и технологической эволюции, которая может помочь в решении проблем ме стных инноваций, миграций и распространения культур- ных особенностей [Wirtz, Lemmen, 2003; Lemmen, Gronenborn, Wirtz, 2011]. Эта модель позволяет дать правдоподобное объяснение возникновению производящего хозяйства во многих регионах мира. В данном исследовании анализ ограничивается Северной Америкой и Западной Евразией. Более подробно будут рассмотрены различия в процессах, приведших к появлению производящего хозяйства в Европе и восточной части Северной Америки, также будет дана приблизительная оценка доколумбового населения североамериканского Вудленда.
Материалы и методы
Модель мирового землепользования и технологической эволюции (Global Land Use and Technological Evolution Simulator – GLUES) [Wirtz, Lemmen, 2003; Lemmen, 2010; Lemmen, Gronenborn, Wirtz, 2011; Lemmen, Wirtz, 2012] описывает развитие региональных социально-культурных особенностей при ограничении процесса биогеографическими условиями. Используемые сообществами ресурсы можно оценить на основе первичной полезной продуктивности региона, которая вычисляется для прошлых эпох с помощью моделируемых аномалий температуры и осадков (системная модель Climber-2) [Claussen et al., 1999] с привлечением базы климатологических данных IIASA [Leemans, Cramer, 1991] и биоклимати-ческой модели предельных объемов растительности [Lieth, 1975]. Концепция модели приводится ниже (подробнее о применяемых алгоритмах и математической основе модели см.: [Wirtz, Lemmen, 2003; Lemmen, Gronenborn, Wirtz, 2011]).
Социально-культурная модель. Социально-культурная сфера характеризуется тремя факторами, описывающими человеческую популяцию. Эволюция каждого фактора во времени проявляет тенденцию приносить все большую пользу связанному с ним населению и, следовательно, способствовать его росту [Dieck-mann, Law, 1996; Wirtz, Eckhardt, 1996]. В данной работе моделируется эволюция распределения каждого фактора в различные моменты, а не единая его величина.
Социально-культурные факторы модели можно кратко охарактеризовать следующим образом.
-
1. Технология – фактор, который описывает экономичность получения продовольствия и эффективность основных видов лечения. В частности, он включает доступно сть инструментов и оружия, а также организацию труда и хранения произведенного продукта. Сюда входит и письменность как инструмент управления и передачи культурного опыта.
-
2. Доля земледелия и скотоводства (с точки зрения энергии, времени и используемой рабочей силы) в общем продовольственном секторе.
-
3. Число земледельческих и скотоводческих форм хозяйствования, доступных для населения региона. Так, например, их четыре в том случае, если в одном сообществе присутствует свиноводство, выращивание ячменя, пшеницы, а также разведение коз. При анализе данного фактора используется работа С. Шеннана [Shennan, 2001], в которой популяционная динамика непосредственно связывается с процессом культурной эволюции.
Параметры модели ограничены требованием успешного моделирования процесса возникновения земледельческих и скотоводческих центров в пяти основных регионах в соответствующие периоды. Согласно исследованию Б.Д. Смита [Smith, 1998, p. 12], это «Плодородный полумесяц» (7500 г. до н.э.), Центральная Мексика (7000 г. до н.э.), Южный (8500 г. до н.э.) и Северный (7800 г. до н.э.) Китай, южная часть Центральных Анд (7000 г. до н.э.) и восточная часть территории США (4500 г. до н.э.)*. В каждой модели со случайным изменением параметров оценка рассчитывается с учетом пространственно-временного расстояния до указанных центров. В данной работе для ретроспективного исследования выбирался набор параметров с самыми лучшими показателями из миллиона имитаций.
Математическая модель основывалась на следующей концепции: для каждого биогеографически определенного региона (со средним размером 127×103 км) в начале моделирования вводятся данные, соответствующие мезолитической популяции охотников и собирателей. Ее численность растет со скоростью, лимитируемой социально-культурными факторами; она ограничивается природными ресурсами. По мере развития технологий ресурсы начинают использовать все более эффективно, и происходят первые случаи окультуривания растений и животных. Рост численности населения и технологически развитое использование природных ресурсов приводит к уменьшению последних. Местный переход к земледелию происходит только тогда, когда сочетание технологий и окультуривания дает больше материальных выгод, чем позволяет образ жизни охотников и собирателей. Как правило, этот процесс завершается менее чем за 200 лет, что хорошо согласуется с эмпирическими данными (см., напр.: [Zvelebil, 1996]).
Производящее хозяйство распространяется из центров своего возникновения в более отдаленные регионы посредством торговли и миграций, механизм которых основывается на концепции различия влияний [Renfrew, Level, 1979], описывающей направление развития и вектор обмена. Миграция происходит со скоростью, пропорциональной длине контактной зоны, поделенной на расстояние между двумя соседями. Влияние торгового обмена соразмерно различию между продуктами торговли. Предполагается, что оно проникает на расстояние в 100 раз большее, чем миграция. Значение торговли и миграции как векторов распространения культурных влияний или расселение народов с последующим принятием новых культурных форм исследовалось в недавней работе [Lemmen, Gronenborn, Wirtz, 2011] с помощью модели GLUES. Было обнаружено, что оба процесса согласуются с радиоуглеродными данными по Европе.
Для различения эмпирически определяемых дат и моделируемых в математической модели была введена шкала моделируемого времени до нашей эры [Ibid.]. В идеале моделируемый год до нашей эры должен быть численно равен календарному.
Биогеографические и биоклиматические факторы. Социально-технологическая модель рассматривалась в географическом контексте в сочетании с данными о флоре прошлых эпох. В модели GLUES растительность оценивается с помощью первичной полезной продуктивности, которая выводится из модели биоклиматического предела [Lieth, 1975] на основе среднегодовых температуры и осадков. Для получения данных о растительности прошлых эпох использовались результаты климатического моделирования переходных условий голоцена средней сложности Climber-2 [Claussen et al., 1999] как аномалии климатологической базы данных IIASA для среднемесячных значений температуры и осадков [Leemans, Cramer, 1991].
На рис. 1 показаны природные ресурсы, связанные с моделируемым переходом к производящему хозяйству. Самая высокая продуктивность растений в 1000 модел. г. до н.э. на юго-востоке Северной Америки, средняя – в восточной и центральной части США и в Европе, низкая – на севере Канады, в районе Скалистых гор и Сахаре (рис. 1, а ). Эти данные репрезентативны для всего моделирования, поскольку изменения климатических параметров настолько незначительны, что величины моделируемой продуктивности не слишком различаются на протяжении всего голоцена. В лесной зоне Европы и восточной части Северной Америки продуктивность растений находится в приемлемом диапазоне (500–1500 г на 1 м2 в год), если преобразовать ее в относительную потенциальную продуктивность земледелия и скотоводства (рис. 1, б ). При функции, определенной [Wirtz, Lemmen, 2003, fig. 1] для перехода от первичной полезной продуктивности к потенциальной продуктивности земледелия и скотоводства, этот потенциал достаточно высок (выше 75 %) и для всей Западной Европы, и для восточной и центральной части США.
Функция перехода от первичной полезной продуктивности к местному видовому разнообразию имеет узкий оптимальный диапазон в 400–600 г на 1 м2 в год, что указывает на наиболее благоприятные условия открытого редколесья для скотоводства и земледелия (однолетние травы, выпас травоядных животных). Моделируемое видовое разнообразие демонстрирует очень высокие показатели по всей Европе, в северо-восточной и центральной части США, но лишь умеренно высокие в юго-восточной (рис. 1, в). Для поправки на ожидаемое более высокое видовое разнообразие на больших участках по сравнению с участками меньшей площади (на что было указано уже почти 200 лет назад [Watson, 1835]) в оценке местного разнообразия подходящих для окультуривания видов на континенте в данном исследовании используется вогнутая зависимость видов и площади [Connor, McCoy, 2001]. Потенциально подходящее для земледелия и скотоводства разнообразие видов в Евразии в 4 раза выше, чем в Северной Америке.
Результаты
Земледелие и скотоводство как основные формы добывания пищи возникают в имитационной модели практически по всему миру к 1490 модел. г. н.э. Охота и собирательство сохраняют свое значение в бореальной зоне Европы и Северной Америки (рис. 2), а также на крупных островах, таких как Исландия (не показано) и Ирландия. На рис. 2 показано моделируемое время перехода к земледелию и скотоводству, когда более половины населения региона обеспечивается пищей с помощью данных форм хозяйствования. Для большей части континентальной Европы это период 7000–4000 модел. гг. до н.э., а для большинства районов восточной части Северной Америки – 2500 модел. г. до н.э. – 1490 модел. г. н.э. Такая разница в моделируемых сроках перехода к производящему хозяйству по сравнению с Европой является следствием различных биогеографических условий на данных субконтинентах [Wirtz, Lemmen, 2003].
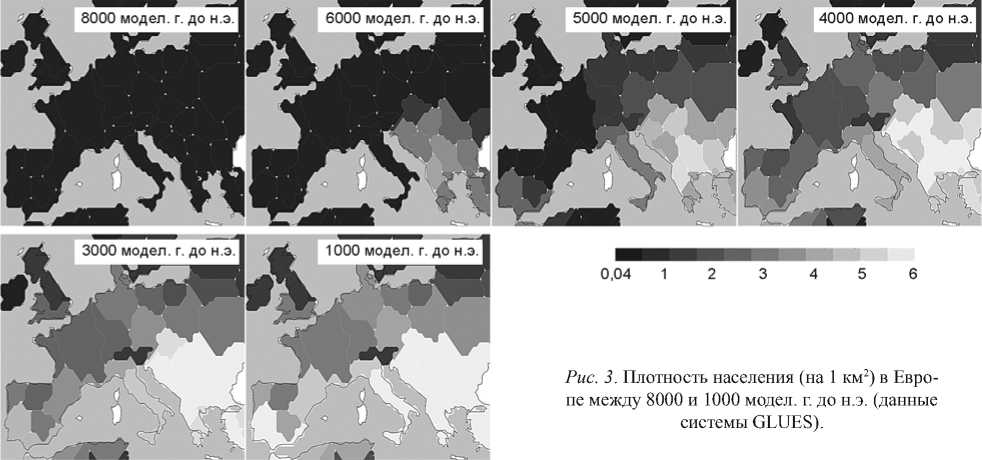
Рост населения в Европе. Модель показывает самое раннее повышение плотности населения ок. 8000 модел. г. до н.э. в Греции, Италии, Анатолии и в северозападной части Пиренейского полуострова (рис. 3). К 6000 модел. г. до н.э. по всей
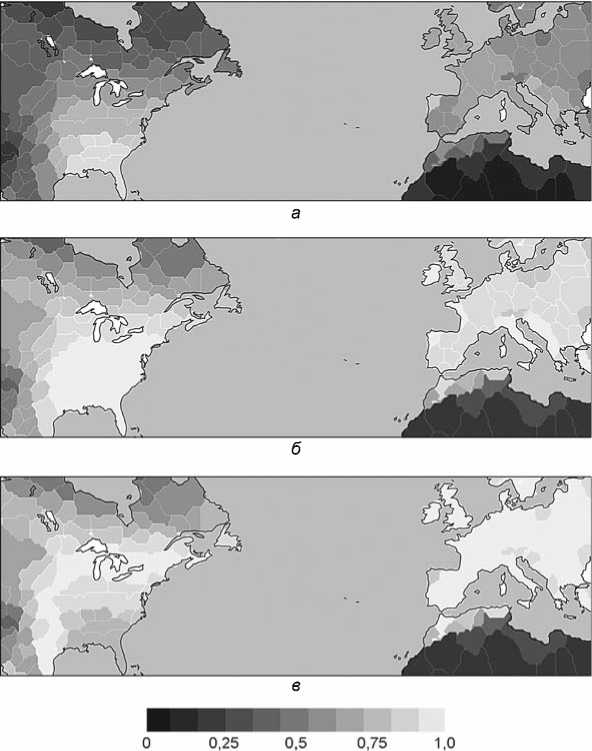
Рис . 1 . Биогеографический контекст для моделирования социально-технологической эволюции в GLUES (биогеографические переменные а – в являются производными от реконструируемой температуры и осадков в 1000 модел. г. до н.э.).
а – первичная полезная продуктивность (килограмм на квадратный метр в год); б – относительная потенциальная продуктивность земледелия и скотоводства; в – относительное число подходящих для окультуривания видов.
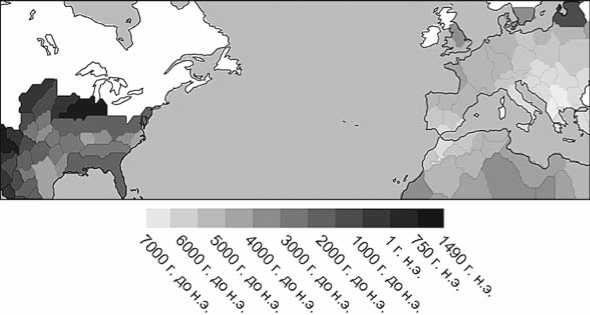
Рис. 2 . Время перехода к земледелию или скотоводству как преобладающему способу добывания пищи, моделируемое с помощью социально-технологической системы GLUES (белый цвет означает, что модель не предсказывает переход в данном месте до 1490 г. н.э.).
4000 модел. г. до н.э.
Рис. 3 . Плотность населения (на 1 км2) в Европе между 8000 и 1000 модел. г. до н.э. (данные системы GLUES).
■■■ 4 5 6
3000 модел. г. до н.э.
1000 модел. г. до н.э.
0,04 1 2 3
6000 модел г до н.э.
5000 модел. г. до н.э.
8000 модел. г. до н.э.
0,04 0,3 0,6 0,9 1,2 1,5
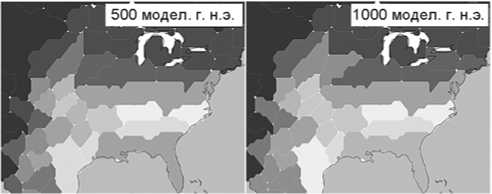
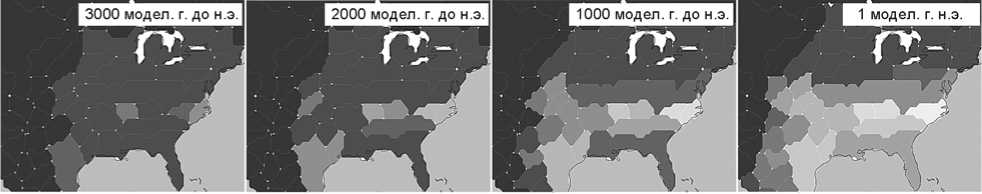
Рис . 4 . Плотность населения (на 1 км2) в Северной Америке между 3000 модел. г. до н.э.
и 1000 модел. г. н.э. (данные системы GLUES).
Европе этот показатель составляет приблизительно 1 чел. на 1 км2. Плотность населения до 3 чел. на 1 км2 локализуется на севере Греции и распространяется на все Балканы и Анатолию. К 5000 модел. г. до н.э. она увеличивается в этих регионах до 5 чел. на 1 км2, возрастает также в Италии и на Пиренейском полуострове. К 4000 модел. г. до н.э. на большей части континентальной Европы плотность населения чуть превышает 2 чел. на 1 км2. Густонаселенными являются Греция, Балканы, Анатолия и прилегающие территории (5–6 чел. на 1 км2). К 3000 модел. г. до н.э. плотность населения в Италии и на юге Пиренейского полуострова повышается до 5 чел. на 1 км2, в то время как на большей части Центральной Европы она достигает уровня чуть менее 3 чел. на 1 км2. К 1000 модел. г. до н.э. вдоль всего Средиземноморского побережья плотность населения составляет
5–6 чел. на 1 км2, а на большей части европейской лесной зоны – почти 3 чел. на 1 км2.
Рост населения в Северной Америке. Здесь моделируемое повышение плотности населения происходит позднее и достигает лишь половины максимальной плотности населения Центральной Европы. Первое увеличение от фонового уровня в 0,3 чел. на 1 км2 проявляется к 3000 модел. г. до н.э. в районе побережья Вирджинии и Северной Каролины (рис. 4)*. Модель показывает самую низкую плотность населения вдоль Скалистых гор (менее 0,1 чел. на 1 км2). К 2000 модел. г. до н.э. плотность населения достигает 0,8 чел. на 1 км2 в этих штатах и несколько превышает фоновый уровень в некоторых областях Восточного Техаса, Арканзаса, Миссури, Теннесси, Миссисипи, Алабамы и Джорджии. К 1000 модел. г. до н.э. она составляет 1 чел. на 1 км2 в южном регионе восточной части Северной Америки и Техасе, а самый высокий показатель наблюдается в районе р. Миссисипи между Арканзасом, Миссури, Кентукки и Теннесси.
С 1 г. н.э. центры сосредоточения населения юговосточной части США сливаются с Восточным Техасом, и возникает крупная область повышенной плотности населения от Мексики до побережья Атлантического океана. Самые высокие показатели в 1,1 чел. на 1 км2 демонстрируют прибрежные штаты Северная Каролина и Вирджиния. К 500 модел. г. н.э. плотность населения в основной области еще увеличивается, а в прибрежных штатах к югу от этой области, на Среднем Западе к западу от нее и на севере вплоть до сороковой параллели достигает 0,6 чел. на 1 км2. К 1000 модел. г. н.э. в восточной части Северной Америки южнее сороковой параллели показатель возрастает до 1,5 чел. на 1 км2 на Атлантическом побережье и в центре, до 0,9 чел. на 1 км2 на остальной территории. В районах к северу от сороковой параллели моделирование показывает низкую плотность населения (до 0,4 чел. на 1 км2 к югу от Великих озер, до 0,35 чел. на 1 км2 в Канаде).
Переход к земледельческому освоению лесной зоны. И для Европы, и для восточной части Северной Америки модель показывает 6500-летний интервал, охватывающий переход к производящему хозяйству – соответственно 7500–1000 модел. гг. до н.э. и 5000 модел. г. до н.э. –1500 модел. г. н.э. (рис. 5). На большинстве европейских территорий переход к производящему хозяйству происходит между 6500 и 4000 модел. гг. до н.э. (рис. 5, а ). В Америке этот процесс начинается после 4000 модел. г. до н.э. и не достигает своего завершения в доколумбовый период, т.е. он оказывается более чем в 2 раза длительнее, чем в Европе (рис. 5, б ), причем на уровне не только субконтинентов, но и отдельных регионов (соответственно ок. 600 и ок. 250 лет).
Между Европой (рис. 5, в) и восточной частью Северной Америки (рис. 5, г) наблюдаются заметные различия в разнообразии форм хозяйственной деятельности. Для Ев- ропы модель показывает постепенное увеличение их числа, соответствующее повышению значения производящего хозяйства. В среднем с 4000 модел. г. до н.э. европейские земледельцы и скотоводы использовали пять различных форм хозяйственной деятельности, и это количество оставалось приблизительно таким же до конца моделируемого периода. Развитие в восточной части Северной Америки началось ок. 4000 модел. г. до н.э. и происходило гораздо медленнее. Число используемых форм хозяйствования постепенно увеличивалось, но не превысило 1,5 в течение всего моделируемого периода. Медленная скорость усвоения новых форм хозяйствования в модели является, в частности, отражением археоботанических процессов, включая длительное время, необходимое для приспособления такого тропического растения, как тео-синте, к субтропическим и умеренным условиям, приемлемым для произрастания кукурузы [Smith, 2011].
Хотя из данной работы и из более ранних исследований очевидно, что различие в максимальных значениях является следствием разных биогеографических условий, следует отметить четкую разницу между двумя субконтинентами по соотношению фактически используемых и потенциально доступных форм хозяйствования на основе местных или ввозимых ре-
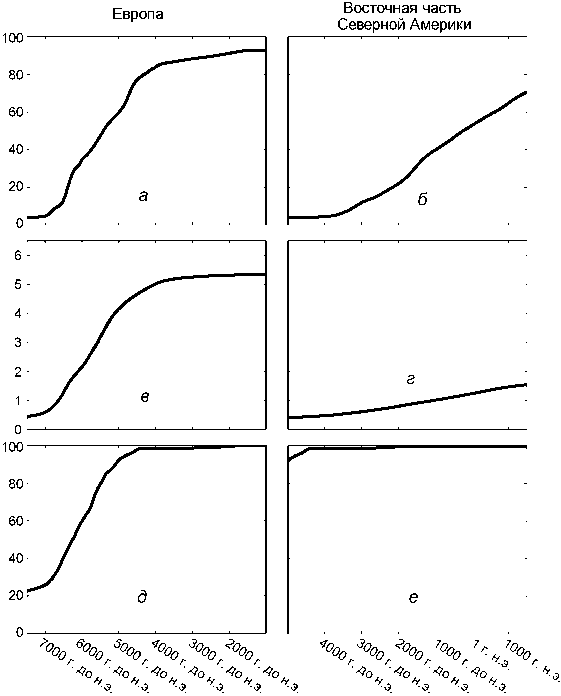
Рис . 5 . График социально-технологического развития в лесной зоне Европы и восточной части Северной Америки (данные системы GLUES усреднены по всем регионам рассматриваемых субконтинентов).
а , б – доля земледелия и скотоводства в системе жизнеобеспечения; в , г – число различных форм хозяйственной деятельности; д , е – процент используемых форм хозяйственной деятельности по отношению к природному потенциалу.
сурсов (рис. 5, д , е ). Доля практиковавшихся видов хозяйственной деятельности в Европе увеличивается с 20 до 100 % к 4000 модел. г. до н.э. Низкая начальная цифра показывает, что все природное богатство практически не использовалось и система испытывала недостаток в человеческих ресурсах: в эпоху неолита не было достаточного количества земледельцев, чтобы освоить весь экономический потенциал. Только с энеолита все разнообразие ресурсов начинает эффективно эксплуатироваться в Европе. В Северной Америке уже в начале перехода к производящему хозяйству (ок. 4000 модел. г. до н.э.) использовался весь спектр экономического потенциала ландшафта. Здесь система ограничивалась природными ресурсами, а проникновение извне новых форм хозяйствования вело к диверсификации хозяйственной деятельности. В археозоологическом контексте такое ограничение ресурсов в первую очередь выражается в отсутствии тягловых животных и внесения удобрений в почву [Piperno, 2011].
Рост численности населения лесной зоны. Средняя плотность населения в Европе (рис. 6) резко возрастает между 7000 и 4000 модел. гг. до н.э. с 0,1 до 3 чел. на 1 км2. Между 4000 и 1000 модел. гг. до н.э. она повышается медленнее, увеличиваясь еще на 1,3 чел.
на 1 км2. На общей площади в 3,1 млн км2 такая плотность населения соответствует истинному размеру популяции в 13 млн чел. к 1000 модел. г. до н.э. Отдельные европейские регионы демонстрируют расхождение в тенденциях роста населения: «взрывной» рост наблюдается в некоторых районах на юго-востоке (например, в Анатолии, Греции, на Балканах), умеренный – в Центральной Европе, очень низкая плотность населения – в северных областях (см. рис. 3).
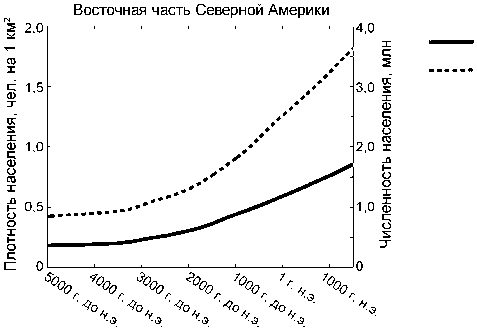
Модель показывает, что численность населения доколумбовой эпохи в североамериканском Вудленде достигала приблизительно 3,5 млн чел. (см. рис. 6). Один миллион из этого числа уже существовал в архаический период; в вудлендский и последующий периоды рост населения составил 2,5 млн. Переход к земледелию и скотоводству не завершился полностью к концу доколумбовой эпохи, и в это время охота и собирательство сохраняли свое значение на половине площади североамериканского Вудленда. Ускоренными темпами этот процесс шел между 4000 модел. г. до н.э. и 1 модел. г. н.э., а в период 1–1490 модел. гг. н.э. его развитие имело линейный характер. Основные особенности перехода к производящему хозяйству на обоих субконтинентах кратко представлены в таблице.
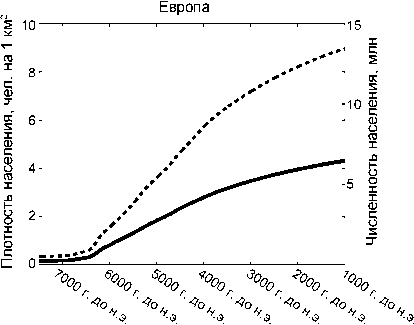
Рис . 6 . График демографического развития в лесной зоне Европы и восточной части Северной Америки (данные системы GLUES усреднены по всем регионам рассматриваемых субконтинентов).
1 – плотность населения; 2 – численность населения.
Основные характеристики моделируемого перехода к земледелию в Европе и восточной части Северной Америки
|
Показатель |
Европа |
Восточная часть Северной Америки |
|
Начало перехода Длительность перехода Потенциал разнообразия местных ресурсов Возможности импорта ресурсов Ограничения раннего периода Пик численности населения |
6500 модел. г. до н.э. 2 500 лет 2,8 2,5 Людские ресурсы 1000 модел. г. до н.э. – 13 млн |
4000 модел. г. до н.э. > 5 500 лет 1,0 0,5 Природные ресурсы 1490 модел. г. н.э. – 3,5 млн |
Обсуждение результатов
«Неолитический пакет» и смешанные формы добывания пищи. Почти мгновенное появление целого ряда разнообразных форм хозяйственной деятельности в Европе может быть связано с крупным «неолитическим пакетом», который, в частности, включал полбу, пшеницу-однозернянку, ячмень; коз, овец и свиней [Willcox, 2005]. Резкое увеличение числа занятых земледелием и скотоводством указывает на появление не только окультуренных видов растений и животных на новых территориях, но и людей, знакомых с земледельческим и скотоводческим образом жизни. Эти процессы можно интерпретировать как выдавливание местного населения – охотников и собирателей переселенцами-земледельцами в соответствии с теорией перенесения культурных форм через миграцию [Ammerman, Cavalli-Sforza, 1973; Sokal, Oden, Wilson, 1991; Haak et al., 2010]. В качестве альтернативного объяснения стремительного перехода к земледелию и скотоводству можно предложить быстрое усвоение новых форм добывания пищи местным населением. Такой механизм подтверждается моделированием в системе GLUES [Lemmen, Gronenborn, Wirtz, 2011]. Авторы работы показывают, что переход к земледелию в Юго-Восточной и Центральной Европе соответствует как переселенческой, так и культурной модели; в обоих случаях важнейшую роль играет усваивание новых культурных форм местным населением.
В североамериканском Вудленде небольшой, но разнообразный набор окультуренных видов состоял из местных растений и аллохтонов при отсутствии одомашненных животных. Локально были доступны киноа, тыква и подсолнечник, входившие в восточный земледельческий комплекс [Delcourt et al., 1998; Smith, 2011]. Постепенно местные культуры дополнились видами, происхождение которых связано с мексиканским центром окультуривания, – кукурузой, бобами и другой разновидностью тыквы. Эти растения часто культивировали совместно как единый комплекс, получивший название «три сестры». Длительное время перехода для всех социально-технологических векторов развития в североамериканском Вудленде указывает на культурную диффузию как основной механизм обмена между регионами, а также на медленное усваивание новых способов добывания пищи [Hart, 1999; Piperno, 2011; Fuller, 2011].
Мировое население и население Америки доколумбового периода. Данные о численности мирового населения, встречающиеся в научной литературе, можно использовать как основу для оценки эффективности модели GLUES в этом отношении. Значения моделируемой численности находятся в широком диапазоне разброса данных из исследований по населению доисторической эпохи (напр.: [Coale, 1974;
McEvedy, Jones, 1978; Biraben, 2003]). На уровне субконтинента оценка в 10 млн жителей Европы, которую дала система GLUES (см. рис. 5, а ), прекрасно согласуется с данными, собранными К. Мак-Эведи и Р. Джонсом [McEvedy, Jones, 1978], или с региональной компиляцией данных Дж. Каплана [Kaplan et al., 2011] для 1000 г. до н.э. После этого, в железном веке, модель GLUES может несколько недооценивать технологическое развитие и рост плотности населения в Евразии, поскольку культурная модель пока еще не учитывает повышение эффективности хозяйствования благодаря обработке металлов и усилению социального расслоения. Для обеих Америк, Африки и Австралии моделирование показывает достоверные результаты вплоть до начала колониальной эпохи, которая также пока не учитывается моделью.
Исследования по численности населения доколумбовой Америки содержат очень разные оценки, иногда различающиеся более чем на порядок: от 10 млн до 150 млн чел. Р. Невл и Д. Берд приводят цифру ок. 60 млн чел. [Nevle, Bird, 2008]. Вероятно, только 10 % из этого числа жили к северу от Мексики, в то время как большая часть населения была сосредоточена в империях ацтеков и инков, а также в бассейне Амазонки [Denevan, 1992]. С учетом приведенных данных очень приблизительная оценка населения американского Вудленда в 4 млн чел. представляется достаточно убедительной.
Лишь недавно [Peros et al., 2010] на основе канадской археологической базы радиоуглеродных данных (CARD) [Morlan] были рассчитаны темпы роста населения в Северной Америке после ледникового периода и вплоть до момента контакта с европейской цивилизацией. Согласно этим расчетам, максимальная численность доколумбового населения составляла 1,8–4,7 млн чел., наиболее вероятно – 2,5 млн чел. Модель GLUES дает величину в 3,5 млн чел. к 1490 модел. г. н.э. для восточной части Северной Америки (см. рис. 5, г ) и 6,5 млн чел. – для всего континента, что представляется вполне реалистичной оценкой.
Критика модели и перспективы. Хорошая согласованность данных социально-культурной модели GLUES с предшествующими исследованиями вызывает некоторое удивление с учетом ее упрощенного подхода и неопределенности многих параметров. Дело в том, что модель детерминированно переводит географическое положение определенного региона и его растительные ресурсы в потенциал для культурной эволюции с универсальной настройкой параметров для всех регионов мира, после чего моделируемые процессы культурного развития в каждом из них и взаимодействие торговли и миграции дают оценки численности населения, продовольственного обеспечения и технологического прогресса. Своим успехом моделирование, скорее всего, обязано тому, что крупномасштабные закономерности культурной эволюции во всем мире основываются на очень похожих функциональных зависимостях от биогеографического контекста и культурных взаимодействий, несмотря на широкий спектр наблюдаемых различий в особенностях культур.
Модель могла бы стать еще более точной, если бы имелись надежные хронологии для большого числа археологических памятников с их одновременной четкой интерпретацией. Для Европы использовались [Lemmen, Gronenborn, Wirtz, 2011] 765 радиоуглеродных дат с неолитической атрибуцией [Pinhasi, Fort, Ammerman, 2005]; модель показала несоответствие данным в среднем в ± 500 лет. Для американского Вудленда привлекалась канадская археологическая база радиоуглеродных данных (CARD) [Morlan]: 3 705 дат из нее имеют обозначение «Woodland», центральные 95 % распределения которых относятся к периоду с 1000 модел. г. до н.э. до 1500 модел. г. н.э. с максимальной частотой в XI в. н.э. В результате применения этих статистических данных к территориям модели GLUES, расположенным в регионе с радиоуглеродными датами вудлендского периода, получены 23 срока появления земледелия и скотоводства в широком диапазоне между 3500 модел. г. до н.э. и 1490 модел. г. н.э. с максимальной частотой в интервале с 2000 модел. г. до н.э. по 1000 модел. г. н.э. Для восточной части Северной Америки моделируемые показатели смещаются к чуть более ранним датам.
Следует отметить, что компиляция радиоуглеродных дат проблематична в связи с различиями в датированных материалах, процедурах калибровки и проверки качества, а также методиках анализа в разных лабораториях и субъективном выборе памятников для включения в базу данных; калибровка сама по себе вносит погрешность (см., напр.: [Stuiver et al., 1998]). Однако еще более серьезную проблему, которую требуется преодолеть при сравнении моделируемых и археологических данных, представляют несоответствия в определении сроков перехода к производящему хозяйству. В модели используется количественный параметр, определяемый как время (модельный год до нашей эры), когда более половины пищи производилось земледельцами и скотоводами. Он не может применяться для археологических данных, где остатки урожая или домашних животных, земледельческие орудия, изображения, свидетельствующие о занятии земледелием или скотоводством, указывают на существование производящего хозяйства, но не на его масштаб. В тех случаях, когда это хозяйство связано с особенностями керамики или каменных орудий, его наличие можно установить и без прямых доказательств. В археологических данных нет ни одного параметра земледельческой и скотоводческой деятельности, который можно было бы напрямую сравнивать с результатами моделирования.
Количественное сравнение можно провести, используя региональные закономерности культурной эволюции по данным радиоуглеродных исследований и моделирования, а также такие косвенные параметры, как плотность или численность населения. При этом самым интересным оказывается отклонение ретроспективных результатов моделирования от исторического пути, что указывает на необходимость объяснения несоответствия с культурной точки зрения. Одним из таких примеров может быть необычно низкая (по отношению к естественным возможностям природной среды) плотность населения сообществ охотников и собирателей в Северной Америке по сравнению с Европой. Это могло заставить модель, в которой данный показатель, детерминированный природными ресурсами, выше, давать более ранний срок возникновения производящего хозяйства в экономике североамериканского Вудленда, чем наблюдается в реальности (см. рис. 4).
Представленная в данной работе модель работает на уровне сообщества. Естественно, в ней не рассматриваются отдельные люди и их культурный выбор, т.е. активное созидание и изменение общества, в котором они живут [Dornan, 2002]. Должна ли подобная модель включать фактор воздействия на коллективном уровне, как это было предложено М. Шенксом и К. Тилли [Shanks, Tilley, 1987]? Я думаю, нет. Моделирование только той части социально-технологической эволюции, которая происходит без прямого воздействия отдельных индивидов, и сравнение результатов с историческими данными позволяют выявить те периоды, когда этот фактор оказывался существенным. Другими словами, прогноз модели следует воспринимать как историческую «нулевую гипотезу», несоответствие моделируемых результатов реальным данным может служить указанием на особые редкие события, имевшие значительные и долгосрочные последствия [Ackland et al., 2007].
Выводы
В данной работе была представлена численная модель регионального перехода к производящему хозяйству. Она способна правдоподобно моделировать потенциальную историю, которую затем можно сопоставить с археологическими данными. Достоверность моделирования подтверждается, например, прогнозированием реальных значений численности населения: 10 млн в Европе к 1000 модел. г. до н.э. и 6,5 млн в Северной Америке (из них 3,5 млн в Вудленде) на 1490 модел. г. н.э.
Несмотря на сходные природные условия американской и европейской лесной зоны и общий аллохтонный характер основных культур, переход к зем- леделию и скотоводству происходил в этих регионах по-разному. Раннее и быстрое принятие нового уклада жизни в Европе, моделируемое в социально-технологической системе GLUES, подтверждается археологическими данными. Модель показывает радиальное распространение земледелия и скотоводства из Восточного Средиземноморья со скоростью, сравнимой с моделью распространения волны, причем все европейское население переходит к производящему хозяйству к 4000 модел. г. до н.э. Переход завершается за достаточно краткий промежуток времени; для него характерен недостаток людских, а не природных ресурсов.
При использовании того же глобального набора параметров для моделирования перехода к производящему хозяйству в американской лесной зоне модель дает иную картину. Здесь охота и собирательство длительно сосуществуют с земледелием и скотоводством; в состав окультуренных видов входят как местные растения, так и аллохтоны; переход на новый уклад жизни осуществляется посредством распространения культурных форм, а не миграции. Этот процесс можно охарактеризовать как ограниченный природными ресурсами. Переход к производящему хозяйству осуществляется в достаточно поздний период, приблизительно с 2000 модел. г. до н.э., так и не получая своего завершения к 1490 модел. г. н.э.
