Регуляция пищевого поведения цыплят при имбалансе лизина и треонина

Автор: Рядчиков В.Г., Тарабрин И.В., Зиганшин Р.Х.
Журнал: Сельскохозяйственная биология @agrobiology
Рубрика: Физиология и биохимия пищеварения
Статья в выпуске: 2 т.42, 2007 года.
Бесплатный доступ
Определяли количество лимитирующих аминокислот в крови, гипоталамусе, печени, мозге у цыплят-бройлеров кросса СК-Русь 2 в связи с их пищевым поведением при содержании на имбалансных по незаменимым аминокислотам рационах. Исследовали влияние перитонеаль-ных инъекций экстрактов головного мозга треонин-имбалансных цыплят и нейропептида Y на пищевое поведение животных в условиях имбаланса рациона по треонину. Изучали наличие у цыплят безусловного рефлекса поиска и идентификации альтернативных источников незаменимых аминокислот при их имбалансе в рационе.
Короткий адрес: https://sciup.org/142133164
IDR: 142133164 | УДК: 636.5.033:636.084.4:591.18.4
Regulation of eating behavior on lysine and threonine imbalance diets in chicks
The authors investigated the eating behavior, growth and content of limitative amino acids in tissues and organs of broiler-chicken of the SK-Rus' 2 cross. Imbalance was made by supplementation of low-protein wheat diet with a crystal amino acid mixture without lysine and threonine, respectively. It was shown that imbalance induced an acute anorectic reaction and the reduction of the growth intensity, concentration of free lysine and threonine in blood plasma, free lysine in hy-pothalamus and also pronounced increase of amino acids content in liver. The same time it was no differences in lysine and threorune content in tissues of liver and brain. Peritoneal injection of brain extracts from threonine imbalanced chicks to the same chicks enhanced the anorectic reaction; neuropeptide Y injection to the same chicks improved the feed consumption. The trail with alternative sources of deficient amino acids demonstrated the animals have a mechanism of identification of the alternative lysine and threonine sources.
Текст научной статьи Регуляция пищевого поведения цыплят при имбалансе лизина и треонина
Определяли количество лимитирующих аминокислот в крови, гипоталамусе, печени, мозге у цыплят-бройлеров кросса СК-Русь 2 в связи с их пищевым поведением при содержании на имбалансных по незаменимым аминокислотам рационах. Исследовали влияние перитонеальных инъекций экстрактов головного мозга треонин-имбалансных цыплят и нейропептида Y на пищевое поведение животных в условиях имбаланса рациона по треонину. Изучали наличие у цыплят безусловного рефлекса поиска и идентификации альтернативных источников незаменимых аминокислот при их имбалансе в рационе.
Признано, что причиной аноректической реакции (анорексия — голодание) животных на имбаланс аминокислот в рационе является дефицит какой-либо незаменимой аминокислоты. В крови дефицит лимитирующих аминокислот распознают сенсорные хеморецепторы, которые находятся в головном мозге. Поскольку содержание лимитирующей аминокислоты наиболее заметно снижается в передней части коры грушевидной доли мозга, несколько меньше — в гипоталамусе, а в остальных 13 из 15 изученных структур головного мозга практически не изменяется, считают, что именно кора грушевидной доли является областью первичной рецепции при анализе дефицита аминокислот. У крыс, которым предлагали имбалансный рацион, инъекция раствора лимитирующей аминокислоты в эту область способствовала повышению потребления корма, а при поражении коры грушевидной доли электрическим током потребление пищи не уменьшалось (1, 2). У лейцин- и треонин-имбалансных крыс низкое содержание этих аминокислот фиксировали в коре грушевидной доли через 30 мин, а отказ от корма несколько раньше — через 12-16 мин (3, 4). В другом эксперименте реакция отвращения к треонин-имбалансной пище наступала через 2 ч, а снижение концентрации треонина в плазме крови — через 3 ч (5). Эти данные позволяют предположить, что первичной областью рецепции при распознавании дефицита или избытка незаменимых аминокислот в организме могут быть и другие структуры, представляющие собой более ранние «системы оповещения», чем рецепторы коры грушевидной доли мозга.
Для растущих животных при имбалансе рациона два фактора оказываются неблагоприятными — дефицит лимитирующей незаменимой аминокислоты и избыток остальных незаменимых аминокислот относительно дефицитной. Имеется много доказательств негативного влияния высокобелкового корма или обогащения рациона избыточным количеством аминокислот на аппетит (6-8). Однако действию фактора избыточности аминокислот в связи с пищевым поведением животных уделяется недостаточно внимания.
В обеспечении работы механизма пищевого поведения задействована нейрогуморальная система. В коре грушевидной доли мозга обнаружены специфические типы молекулярных рецепторов для нескольких трансмиттерных систем, которые вовлечены в реакцию на дефицит незаменимых аминокислот. В том числе это норэпинефрин, γ -аминомасляная кислота, серотонин, дофамин, оксид азота, модулирующие работу глутаминергических синапсов в коре грушевидной доли (9). Эти системы осуществляют проведение сигнала и активируют последующие реакции деградации и синтеза белка при изменении генной активности клеток мозга (7, 10, 11). Имеется также ряд исследований, показывающих роль нейропептидов и гормонов в регуляции пищевого поведения (12). В частности, инъекция нейропептида Y в количестве 1-1,5 нмоль в кору грушевидной доли мозга треонин-имбалансных крыс вызывала снижение потребления корма. Соматостатин имел двойное действие: при инъекции 1 пмоль аппетит улучшался, при инъекции 2 пмоль ухудшался. У цыплят на сбалансированном рационе нейропептид Y стимулировал потребление корма (13). Гормон, ингибирующий гонадотропин, при интрацеребровентрикулярной инъекции цыплятам повышал аппетит, антисыворотка к этому гормону вызывала аноректическую реакцию. Однако пока нет сведений об участии нейропептидов в специфических реакциях на дефицит какой-либо незаменимой аминокислоты или избыточное количество недефицитных аминокислот (14). Поскольку система механизмов пищевого поведения формируется в головном мозге на молекулярном уровне, на первом этапе исследования представляло интерес получить экстракты мозга и проверить их действие на пищевое поведение цыплят после интраперитонеальной инъекции.
Цель нашей работы состояла в следующем: изучить эффективность потребления корма цыплятами-бройлерами в условиях имбаланса лизина и треонина в рационе; оценить влияние разных форм баланса аминокислот в рационе на концентрацию свободных аминокислот в крови и их содержание в печени, мозге, гипоталамусе; исследовать способность животных к поиску и идентификации альтернативных источников дефицитной аминокислоты, а 43
также к адаптации в условиях имбаланса аминокислот; сравнить действие интраперитонеальной инъекции нейропептида Y и экстрактов мозга имба-лансных особей на пищевое поведение цыплят.
Методика . О п ы т 1 . И з у ч е н и е м еха н и з м о в р е гул я ц и и пищевого поведения цыплят в условиях имбаланса лиз и н а и т ре о н и н а в р а ц и о н е . Опыт проводили на четырех группах цыплят-бройлеров кросса СК-Русь 2. Группы по 12 особей подбирали методом аналогов среди цыплят в возрасте 18 сут. Протоколы работы с животными утверждены Ученым советом зооинженерного факультета Кубанского государственного аграрного университета.
Перед началом эксперимента цыплят всех групп в течение 3 сут кормили одним и тем же комбикормом согласно нормам ВНИТИП-2003 (15). После этого I группу (контроль) перевели на основной рацион (табл. 1), который включал зерно пшеницы, смесь аминокислот (первой лимитирующей аминокислотой в рационе был лизин, второй — треонин), добавки витаминов, макро- и микроэлементов по нормам ВНИТИП-2003. Кроме того, в корм вводили муку из пшеничной соломы (доля по массе 3 %) для обеспечения животных необходимым количеством клетчатки, а также препарат β -глюканазного фермента Роксозим G («Ф. Хофманн-Ля Рош Лтд.», Швейцария), который улучшает переваримость некрахмалистых полисахаридов. В IV группе (опыт, скорректированный рацион) основной рацион обогащали всеми недостающими незаменимыми и заменимыми L-аминокислотами, в том числе лизином и треонином, до 100 % потребности. Аминокислоты («Аджиномото Ко Лтд.», Япония) вносили в виде кристаллических препаратов. Добавки (г/кг корма) составили: лизин·HCl (98 %) — 10,5; DL-метионин — 3,0; цистин — 3,0; треонин (98 %) — 5,8; триптофан — 1,3; аргинин HCl·H 2 O — 9,5; глицин + серин — 2,2; гистидин·HCl — 3,4; изолейцин — 6,2; лейцин — 10,4; фенилаланин + тирозин — 7,6; валин — 6,1; глутаминовая кислота — 18,0; пролин — 10,0; аланин — 10,0; аспарагин — 18,0. Цыплята II группы (опыт, имбаланс лизина) получали основной рацион плюс смесь кристаллических препаратов всех недостающих незаменимых и заменимых аминокислот до 100 % потребности, кроме лизина, количество которого оставалось таким же, как в I группе (30 % потребности). Аналогично III группа (опыт, имбаланс треонина) получала основной рацион, который был обогащен всеми недостающими аминокислотами, за исключением треонина.
1. Содержание компонентов в основном рационе цыплят-бройлеров кросса СК-Русь 2 при изучении пищевого поведения (опыты 1-4)
|
Компонент |
Доля по массе, % |
и треонина по рационам со- |
|
Пшеница |
90,8 |
ставили соответственно 10,45; |
|
Масло подсолнечное |
2,5 |
0,30; 0,305 (основной рацион); |
|
Трикальцийфосфат |
3,0 |
18,4; 0,28; 8,5 (лизин- |
|
Мука из пшеничной соломы |
3,0 |
|
|
Премикс микроэлементный Веталлас Евровед |
0,1 |
имбалансный рацион); 18,6; |
|
Премикс витаминный Веталлас Евровед |
0,1 |
1,12; 0,290 (треонин- |
|
Антиоксидант |
0,015 |
|
|
Флавомицин |
0,006 |
имбалансный рацион); 18,9; |
|
Роксозим G-2 |
0,012 |
1,12; 8,5 (скорректированнный |
|
NaCl |
0,4 |
|
|
Смесь незаменимых аминокислот: |
рацион). Цитрат натрия вво- |
|
|
лизин |
0,30 |
дился в рационы II-IV групп |
|
метионин + цистин |
0,41 |
|
|
треонин |
0,305 |
для оптимизации соотношения |
|
триптофан |
0,12 |
натрия и хлора, который по- |
|
аргинин |
0,50 0,89 |
ступал с препаратами лизина, |
|
глицин + серин |
||
|
гистидин |
0,23 |
аргинина и гистидина. При |
|
изолейцин |
0,38 |
этом NaCl из рационов II-IV |
|
лейцин |
0,68 |
|
|
фенилаланин + тирозин |
0,15 |
групп исключали. Кормили цы- |
|
валин |
0,46 |
плят вволю в течение 11 сут, |
Показатели содержания (доля по массе, %) сырого протеина (белок пшеницы +
П р и м е ч а н и е. Характеристики пищевой ценности рациона: обменная энергия — 320,0 ккал/100 г, содержание сырого протеина — 10,45 %, содержание сырой клетчатки — 3,2 %, Ca — 1,0 %, P общий — 0,68 %, P доступный — 0,42 %, Na — 0,18 %, Cl — 0,24 %.
рост учитывали индивидуально, потребление корма — по группам.
Для определения сво- бодных аминокислот в плазме крови, печени и мозге в конце опыта четырех
цыплят из каждой группы умерщвляли методом декапитации через 2 ч после начала потребления корма. Головы сразу же замораживали в жидком азоте, затем в течение минуты препарировали образцы целого мозга и отдельно гипоталамуса. Образцы хранили в сосуде Дьюара в жидком азоте (-196 oC). Пробы крови и печени отбирали при убое цыплят. Кровь гепаринизировали, форменные элементы отделяли центрифугированием, после чего плазму замораживали при –16 оС. Образцы печени сразу после изъятия замораживали в жидком азоте.
При анализе плазмы крови к 300 мкл исходного материала добавляли 33 мкл 44 % сульфосалициловой кислоты для осаждения белков. В течение 10 мин тщательно перемешивали содержимое пробирок на электромешалке, 45
после чего центрифугировали в течение 3 мин (320 g ). Отбирали по 30 мкл супернатанта, добавляли к отобранному образцу 270 мкл 0,1 н. HCl, перемешивали. При подготовке проб печени, мозга и гипоталамуса для аминокислотного анализа навески ткани по 10 мг гомогенизировали в 180 мкл дистиллированной воды. Осаждение белка и центрифугирование проводили так же, как для образцов крови. Содержание аминокислот в пробах определяли на анализаторе Toshiba 835 (Япония).
Опыт 2. Способность цыплят-бройлеров к поиску и идентификации альтернативных источников лизина и треон и н а п ри и м бал ан се ра ц и о н а . Методом аналогов сформировали две группы цыплят-бройлеров кросса СК-Русь 2 в возрасте 20 сут по 10 гол. в каждой. Цыплятам из I группы предлагали рацион с имбалансом лизина, из II — c имбалансом треонина. Рационы составляли так же, как в опыте 1. На этих рационах птицу содержали 8 сут, потребление воды не ограничивали. В следующие 4 сут птице из I группы (имбаланс треонина) ставили две поилки — c водой и с 0,5 % водным раствором треонина. Во II группе (имба-ланс лизина) цыплятам предлагали поилку с водой и поилку с 1 % водным раствором гидрохлорида лизина. Затем на 4-е сут поилки с раствором аминокислоты убирали, оставляя только поилки с чистой водой. Потребление воды, водных растворов аминокислот и корма учитывали посуточно.
Опыт 3. Влияние имбаланса на поведенческие реакции цыплят (поедание корма, передвижение по клетке , со н , п от ребл е н и е вод ы ) . Методом аналогов сформировали две группы цыплят-бройлеров кросса СК-Русь 2 в возрасте 20 сут по 10 гол. в каждой. Условия содержания и кормления цыплят были такими же, как в опытах 1 и 2. В течение первых 5 сут обеим группам предлагали скорректированный рацион (см. методику опыта 1). Далее I группа продолжала получать тот же рацион, а цыплят II группы перевели на корм с имбалансом треонина. С этого момента проводили поминутный хронометраж поведения животных в течение первых 6 ч светового дня. Через 3 сут хронометраж повторили.
Опыт 4. Действие интраперитонеальной инъекции э кст р а кт о в г ол о в н о г о м о з г а и н е й р о п е п т и д а Y ( N PY ) н а п и ще в ое п о веде н и е ц ы п ля т - б ро йл е ро в . Экстракты головного мозга для инъекций получали от цыплят из III (имбаланс треонина) и IV группы (скорректированный рацион) опыта 1. Для изучения действия нейропептида Y (свиного) использовали коммерческий препарат ICN 2004, Neuropeptide Y (82785-45-3).
На первом этапе экстракции головной мозг от четырех особей объединяли в одну пробу, добавляли 1 М уксусную кислоту из расчета 10 мл на 1 г ткани и гомогенизировали на льду. Для предотвращения возможных ферментативных процессов перед гомогенизацией в образцы вводили ингибиторы протеаз: фенилметилсульфонилфторид (PMSF, 10-4 моль, ингибитор сериновых и цистиновых протеаз), pepstatin A (10-6 моль, ингибитор кислых протеаз), этилендиаминтетрауксусную кислоту (ЭДТА, 10-3 моль, ингибитор протеаз, содержащих двухвалентные металлы). Использовали гомогенизатор Polytron PT-3000 («Kinematica AG», Швейцария, 45 000 об/мин). Гомогенат центрифугировали в течение 10 мин при 2200 g на центрифуге Eppendorf (Германия) при температуре +2 оС. Полученный супернатант и осадок замораживали при –80 оС и лиофилизировали. На втором этапе экстрации лиофилизированный супернатант перерастворяли в 1 М уксусной кислоте, повторно центрифугировали для более полной очистки от нерастворимых остатков и гидрофобных белков и лиофилизировали.
В эксперимент по интраперитонеальной инъекции отобрали четыре группы по 10 цыплят-бройлеров кросса СК-Русь 2 в возрасте 21 сут с живой массой 380 г. Условия содержания и кормления птицы были такими же, как в опыте 1. Все группы потребляли рацион с имбалансом треонина (см. методику опыта 1). Цыплятам I группы (контрольной) вводили по 1 мл физиологического раствора. Животные II группы получали интраперитонеально нейропептид Y из расчета 90 мкг/кг живой массы (35 мкг в 2 мл физиологического раствора каждому цыпленку). В III группе животным однократно вводили экстракт головного мозга от цыплят, содержавшихся на скорректированном рационе, а в IV группе — экстракт мозга от треонин-имбалансных цыплят из расчета 120 мг/кг живой массы (45 мг в 2 мл физиологического раствора). За 1 ч до инъекций начали вести поминутный хронометраж пищевого поведения. Потребление корма учитывали по группам.
Результаты . О п ы т 1 . И зуч е н и е м еха н и з м о в р е гул я ц и и пищевого поведения цыплят в условиях имбаланса лиз и н а и т ре о н и н а в ра ц и о н е . Цыплята I группы, которых содержали на основном низкобелковом рационе, довольно хорошо поедали корм (табл. 2). Во II и III группах обогащение рационов смесями аминокислот (соответственно без лизина и треонина) вызывало резкое снижение потребления корма, хотя содержание лизина и треонина в рационах указанных групп оставалось практически таким же, как в I группе (см. табл. 2). Эти данные показывают, что фактором, вызывающим снижение пищевой активности при имбалансе рациона, явля-47
ется избыток других аминокислот относительно лимитирующих — лизина и треонина. У животных, которых содержали на скорректированном по аминокислотам рационе (100 % нормы, включая лизин и треонин), потребление корма было самым высоким.
Интенсивность роста бройлеров напрямую зависела от количества потребленного корма. Максимальный среднесуточный прирост живой массы отмечали у цыплят на скорректированном рационе (IV группа), наименьший — при имбалансе лизина (II группа). В III группе (имбаланс треонина) уменьшение живой массы цыплят в среднем за сутки составляло 3,1 г. Цыплята, получавшие основной низкобелковый пшеничный рацион, относительно неплохо росли и выглядели вполне здоровыми. В ранее проведенных экспериментах мы не наблюдали повышения потребления корма животными как способа адаптации к имбалансу рациона (16).
2. Рост и потребление корма по группам (I-IV) цыплят-бройлеров кросса СК-Русь 2 при содержании на рационах с разным балансом незаменимых аминокислот
|
Показатель |
I (основной рацион) |
II (имбаланс лизина) |
III (имбаланс треонина) |
IV (скорректированный рацион) |
|
Живая масса, г: в начале опыта |
679,5±22,3 |
675,4±21,7 |
664,2±20,2 |
670,9±30,0 |
|
в конце опыта |
836,9±25,1 |
689,6±29,9 |
630,1±17,7 |
1278,7±54,6 |
|
Среднесуточный прирост |
14,3±1,1 |
1,2±0,6 |
–3,1±1,2 |
49,8±0,9 |
|
живой массы, г Потребление, ½ (гол ˛ сут)-1: корм, г |
88,3 |
44,4 |
38,9 |
98,2 |
|
белок пшеницы + аминокислоты, г |
9,22 |
8,15 |
7,22 |
18,85 |
|
лизин, мг |
265 |
124 |
436 |
1100 |
|
треонин, мг |
260 |
377 |
109 |
840 |
|
П р и м е ч а н и е. Состав рационов см. в разделе «Методика» . |
||||
Концентрация свободных аминокислот в плазме крови цыплят. У цыплят, получавших имбалансные рационы с низким содержанием лизина и треонина на фоне обогащения нелимитирующими аминокислотами (II и III группы), при снижении потребления корма происходило достоверное уменьшение концентрации соответствующих дефицитных аминокислот в плазме крови (Р < 0,05 и Р < 0,001) по сравнению с этим показателем у животных в контрольной группе (табл. 3).
В плазме крови лизин-имбалансных цыплят концентрация пролина, аланина, гистидина, триптофана была достоверно ниже, а глицина, орнитина и аргинина — выше по сравнению с их концентрацией в плазме крови цыплят, содержащихся на основном рационе. У треонин-имбалансных цыплят низкой была концентрация оксипролина, аспарагиновой и глутаминовой кислот, гистидина и триптофана, но существенно увеличивалась концентрация глицина.
3. Количество свободных аминокислот (мг%) в тканях цыплят-бройлеров кросса СК-Русь 2 по группам (I-IV) при содержании на рационах с разным балансом незаменимых аминокислот ( n = 4)
|
Аминокислота, метаболит |
I (основной рацион) |
II (имбаланс лизина) |
III (имбаланс треонина) |
IV (скорректированный рацион) |
|
Плазма крови |
||||
|
Таурин |
1,10±0,19 |
0,97±0,18 |
1,96±0,14*** |
1,18±0,14 |
|
Мочевина |
0,14±0,04 |
0,10±0,01 |
0,30±0,09 |
0,12±0,01 |
|
Треонин |
10,13±0,58 |
19,78±0,66*** |
6,76±0,59*** |
16,39±1,34*** |
|
Лизин |
1,65±0,12 |
0,98±0,09* |
7,15±0,77** |
3,00±0,04** |
|
Сумма аминокислот |
71,0±2,0 |
81,1±3,9* |
69,2±3,1 |
69,8±3,9 |
|
Печень |
||||
|
Таурин |
33,53±5,76 |
54,04±0,31** |
76,19±9,51*** |
71,06±2,75*** |
|
Мочевина |
3,65±0,50 |
5,59±0,041** |
9,18±1,74** |
5,77±1,20 |
|
Треонин |
93,97±9,05 |
128,63±0,60** |
92,3±2,98 |
129,82±5,96*** |
|
Лизин |
66,23±3,36 |
70,61±0,21 |
71,93±2,05 |
106,73±11,70*** |
|
Сумма аминокислот |
1189,3±64,8 |
1334,3±65,9 |
1464,6±71,9** |
1699±80,4*** |
|
Мозг |
||||
|
Таурин |
32,03±0,75 |
36,78±0,20 |
37,78±2,00 |
42,03±3,75 |
|
Мочевина |
2,74±0,10 |
3,33±0,01* |
3,16±0,31 |
3,04±0,15 |
|
Треонин |
52,64±3,57 |
72,65±0,42* |
59,07±2,38 |
77,89±2,26** |
|
Лизин |
17,84±1,46 |
19,44±0,13 |
22,66±2,63 |
21,05±0,73 |
|
Сумма аминокислот |
684,7±3,6 |
709,5±31,1 |
692,6±55,5 |
745,5±40,7 |
|
Гипоталамус |
||||
|
Таурин |
17,89±0,13 |
22,64±0,16 |
13,64±1,38 |
31,28±0,13*** |
|
Мочевина |
2,00±0,00 |
2,26±0,00 |
1,58±0,05** |
2,52±0,04** |
|
Треонин |
39,9±2,14 |
53,12±0,39 |
38,35±3,22 |
67,65±4,29** |
|
Лизин |
15,5±0,88 |
11,46±0,02* |
13,03±0,23 |
20,91±2,49 |
|
Сумма аминокислот |
556,0±18,0 |
582,0±12,1 |
404,8±12,1** |
623,1±9,8* |
|
* P < 0,05; ** P < 0,01; *** P < 0,001 (достоверность различий относительно I группы). |
||||
Суммарное количество свободных аминокислот в плазме крови цыплят на рационе с имбалансом лизина было существенно выше по сравнению с этим показателем у животных остальных групп, причем в основном за счет двух аминокислот — треонина и глицина. Ранее в опытах на белых крысах, получавших рацион с дефицитом лизина, мы также наблюдали обратную зависимость между концентрацией лизина и треонина в плазме крови (17).
Содержание свободных аминокислот в печени. Известно, что печень является органом, в котором осуществляется переаминирова- ние аминокислот и переработка аммиака, освобождаемого при дезаминировании аминокислот, в мочевину. Следует отметить тот факт, что содержание лизина и треонина в печени в группах с имбалансом рациона оказалось на уровне контроля (основной рацион), хотя потребление этих аминокислот в опыте снижалось более чем в 2 раза относительно контроля. Вместе с тем общее содержание аминокислот у имбалансных цыплят было значительно выше, чем у цыплят, получавших основной рацион (соответственно на 12,2 и 23,1 %). Задержка значительного количества аминокислот в печени, по-видимому, обусловлена трудностями быстрой утилизации избыточных потоков нелимитирующих аминокислот при дефиците лизина и треонина. О напряженности этих процессов свидетельствует также высокое содержание мочевины и таурина в печени цыплят имбалансных групп.
В печени цыплят, получавших скорректированный рацион, суммарное содержание свободных аминокислот тоже было весьма значительным — соответственно на 43, 27 и 16 % выше, чем у животных из I, II и III групп. Однако с учетом того, что количество потребленного белка (белок пшеницы + препараты аминокислот) в IV группе составило соответственно 208, 233 и 265 % относительно его количества, полученного цыплятами из I, II и III групп, приведенные значения суммарного содержания аминокислот представляются вполне нормальными. В данном случае существенную долю в увеличение общего показателя вносили заменимые аминокислоты — глутаминовая, аспарагиновая, глицин, а из незаменимых — лизин.
Содержание свободных аминокислот в мозге. Следует отметить, что в мозге цыплят разных групп содержание как отдельных аминокислот, так и их общая сумма оказались практически одинаковыми, несмотря на огромные различия в потреблении аминокислот с пищей (исключение составил треонин, содержание которого в материале от лизин-имбалансных цыплят и цыплят из группы, получавшей скорректированный рацион, было достоверно выше — соответственно P < 0,05 и P < 0,01). Более того, в мозге цыплят из II и III групп количество дефицитных лизина и треонина не снижалось относительно контрольной группы, которая получала основной рацион. Эти данные могут быть доказательством важной роли гематоэнцефалического барьера в регуляции содержания свободных аминокислот на уровне, соответствующем гомеостатическим параметрам мозга как наиболее важного органа. Известно, что в капиллярно-транспортной системе аминокислоты конкурируют за гематоэнцефалический барьер (18). Возможным механизмом регу- ляции конкурентных потоков является поддержание гематоэнцефалическим барьером аминостатического гомеостаза.
Содержание свободных аминокислот в гипоталамусе. В гипоталамусе цыплят из группы с имбалансом лизина его содержание было достоверно ниже (P < 0,05), чем у контрольных цыплят (основной рацион). Однако в случае с треонином (цыплята из III группы в сравнении с I) соответствующую разницу мы не выявили. В то же время суммарное содержание аминокислот у особей в III группе было намного ниже, чем в остальных, что неудивительно, так как цыплята в этой группе очень плохо поедали корм и теряли живую массу, особенно во второй половине опыта.
Можно полагать, что в гипоталамусе гематоэнцефалический барьер не сортирует потоки аминокислот по гомеостатическому принципу. По-видимому, в данном случае функция гипоталамуса — анализ состава доставляемых кровью аминокислот per se, который необходим для активации системы нейромедиаторов и нейропептидов, участвующих в формировании соответствующего пищевого поведения.
ис
Опыт 2. Способность цыплят-бройлеров к поиску и идентификации альтернативных источников лизина и треон и н а п р и их и м бала н се в ра ц ио н е . Предоставление недостающих аминокислот в виде водных растворов лизина или треонина в дополнительной поилке выявило способность цыплят быстро находить альтернативные источники дефицитного пищевого компонента и пить в значительно большем количестве не воду, а раствор недостающей аминокислоты (рис.). При перемене местоположения поилок животные быстро находили источник аминокислоты, хотя сами поилки не различались по форме, а вода и водный раствор аминокислот — по цвету и запаху. С повышением потребления растворов недостающих аминокислот потребление корма в группах на имба-лансных по лизину и треонину рационах увеличилось на 60-70 %, что, в свою очередь, способствовало повышению среднесуточных приростов живой массы соответственно с 2,4 до 31,2 г и с 11,7 до 45 г. После 4-суточного периода с двумя поилками цыплятам снова оставляли одну поилку с водой. Это вызвало резкое снижение потребления корма и среднесуточных приростов живой массы (соответственно до 2,6 и 19,2 г).
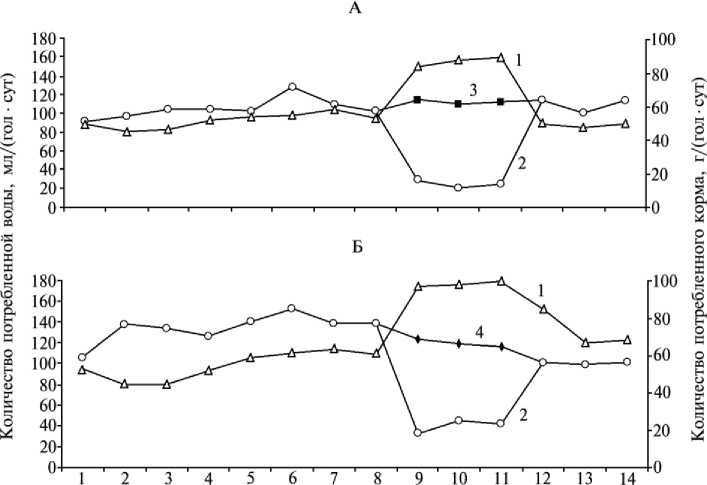
Время, сут
Потребление корма (1), воды (2), водных растворов лизина (3) и треонина (4) цыплятами-бройлерами кросса СК-Русь 2 при имбалансе лизина (А) и треонина (Б) в рационе.
Результаты этих опытов свидетельствует о том, что животные обладают очень чувствительным механизмом контроля аминокислотного состава пищи, рефлексом поиска и идентификации альтернативных источников дефицитных аминокислот.
Опыт 3. Влияние имбаланса рациона на поведен-чес кие реакции цыплят (поедание корма, передвижение по клетке, сон, потребление воды). Хронометраж поведенческих реакций показал, что со 2-го ч после перевода цыплят на рацион с имба-лансом треонина продолжительность потребления корма резко сократилась. Через 3 ч доля времени, затрачиваемого цыплятами на перемещение по клетке и потребление воды возросла соответственно на 40 и 32 %, на сон — уменьшилась на 16 %. Через 3 сут продолжительность потребления корма увеличилась, затраты времени на перемещение по клетке возросли в 3 раза (видимо, вследствие поиска альтернативного источника пищи), время, в течение которого животные пили воду, сократилось. В контрольной группе, которую содержали на скорректированном рационе, животные 65-75 % времени спали или лежали; треонин-имбалансные цыплята на сон затрачивали 40-55 % времени наблюдения. Результаты оценки поведенческих реакций цыплят (табл. 4) свидетельствуют о более возбужденном состоянии центральной нервной системы при несбалансированном по аминокислотам питании и, по-видимому, о более высоких затратах энергии на физическую активность.
Опыт 4. Действие интраперитонеальной инъекции экстрактов головного мозга и нейропептида Y (NPY ) на пищевое поведение цыплят - бройлеров. Динамика потребления треонин-имбалансными цыплятами (п = 10) корма до и после интраперитонеальной инъекции была следующей: в контроле (I группа опыта 4, инъекция физиологического раствора) — 4,0; 3,0; 3,0 (соответственно 1 ч до инъекции, 1-й и 2-й ч после инъекции); в варианте с NPY (II группа опыта 4) — Ж 4,0; 2,0; 4,0; при введении материала от цыплят из группы со скорректированным рационом (III группа опыта 4) —Ж 4,0; 3,0; 3,0; при инъекции экстракта мозга треонин-имбалансных цыплят (Ж^ группа опыта 4) — 4,0; 2,0; 2,0 г/гол. Общее потребление корма за сутки в I, II, III и IV группах составило соответственно 45,0; 49,0; 42,0 и 38,0 г/гол.
4. Поведенческие характеристики цыплят-бройлеров кросса СК-Русь 2 при содержании на скорректированном и треонин-имбалансном рационах (% времени наблюдения, п = 10)
|
Группа |
Пищевая активность |
Перемещение по клетке |
Сон, отдых |
Потребление воды |
|
I (скорректированный рацион, |
11,3±1,67 |
11,8±4,50 |
73,3±7,33 |
3,5±1,17 |
|
контроль) |
||||
|
II (имбаланс треонина) |
8,0±3,00 |
36,3±2,67* |
54,3±6,00 |
1,3±0,33 |
* P < 0,05.
П р и м е ч а н и е. Условия опыта см. в разделе «Методика».
Таким образом, в 1-й ч после инъекции нейропептида Y и экстрактов мозга во всех группах отмечали снижение потребления корма. За 2-й ч в группе, которой инъецировали нейропептид Y, потребление корма восстановилось до уровня почасового потребления перед инъекцией. Суточное потребление корма в этой группе оказалось максимальным. Цыплята, инъецированные экстрактами мозга животных из групп, содержавшихся на имба-лансном и скорректированном рационах (соответственно III и IV группы опыта 1), в течение первых 2 ч не могли восстановить потребление корма до исходного уровня. Особенно это было заметно в группе, которой инъецировали экстракт мозга треонин-имбалансных цыплят: в целом за сутки оно отставало на 16 % от контроля и на 22,5 % — от потребления в группе, получившей инъекцию нейропептида Y. Эти данные могут свидетельствовать о наличии в экстрактах мозга треонин-имбалансных цыплят веществ, активно действующих на пищевое поведение, что дает основание продолжать исследования 53
молекулярных механизмов его регуляции при разных формах баланса амино- кислот.
Итак, в наших опытах сбалансированные дозы аминокислотных добавок к основному низкобелковому рациону использовались цыплятами и стимулировали их рост. В то же время в имбалансных рационах из-за некомпен-сированности по лизину и треонину полное обеспечение потребностей животных в остальных аминокислотах оказалось «излишним». Животные не могли использовать эти аминокислоты для синтеза белка и роста, вследствие чего на рационах с имбалансом потоки аминокислот, которые не утилизировались по назначению, и их метаболитов (аммиака, мочевины, таурина) возрастали, причем в большей степени, чем на низкобелковом основном рационе.
Имеются данные, что при увеличении содержания белка или аминокислот в условиях имбаланса наблюдается повышение активности ферментов деградации дефицитных аминокислот. Например, активность треониновой дегидратазы в большей мере повышалась в ответ на увеличение количества белка, а не треонина (19). Добавки смеси из 17 кристаллических аминокислот в полусинтетический рацион без треонина вызывали рост активности треониновой дегидратазы в печени цыплят более чем в 3 раза (20). У крыс на гистидин-имбалансном рационе наблюдали подобную реакцию: активность фермента деградации гистидина —Ж гистидазы и содержание ее мРНК в печени значительно возрастали при введении в казеиновый рацион смеси аминокислот без гистидина (21), а при добавлении к кукурузному рациону смеси аминокислот без лизина активность фермента лизин- а -кетоглутаратредуктазы увеличилась в 2 раза (22).
Повышение активности ферментов деградации лимитирующих аминокислот в ответ на увеличение общего количества белка или аминокислот кажется крайне нелогичным, поскольку дефицит лимитирующей аминокислоты усугубляется, аноректическая реакция на имбаланс становится более острой по сравнению с низкобелковым рационом. Объяснить подобный феномен трудно, если не принять во внимание тот факт, что в процессе эволюции механизм регуляции активности ферментов деградации формировался в естественной среде обитания животных. В природе не встречаются источники пищи с крайними отклонениями по содержанию той или другой незаменимой аминокислоты либо ее полным отсутствием. Такой дисбаланс создается в искусственных рационах, приготовленных с использованием химических препаратов аминокислот. Поэтому активность ферментов деградации регулируется не 54
столько субстратом — целевой аминокислотой, сколько общим количеством белка (аминокислот).
Пул свободных аминокислот в жидкостях и тканях формируется за счет экзогенных аминокислот, поступающих в результате переваривания пищи, и эндогенных, образовавшихся в результате гидролиза клеточными протеазами белков организма в процессе их обновления. Часть эндогенных аминокислот окисляется с образованием аммиака и мочевины, другая часть участвует в ресинтезе белков. Разделить содержащиеся в плазме крови, печени и мозге свободные аминокислоты по происхождению невозможно. Видимо, вследствие больших эндогенных потоков в органах с наименьшим периодом полужизни белков, какими являются печень и мозг, мы не обнаружили низкого содержания лизина и треонина при соответствующих имбалансах. В плазме крови цыплят наблюдалось достоверное снижение концентрации этих аминокислот через 2 ч после начала кормления. В гипоталамусе снижалось содержание лизина, а содержание треонина оставалось таким же, как у цыплят на основном рационе.
Причиной анорексии при имбалансе может быть не только низкий уровень дефицитной, но и избыток остальных аминокислот относительно лимитирующей. Мы отмечали существенное увеличение содержания суммы аминокислот, мочевины и таурина в печени имбалансных цыплят по сравнению с контролем (при том, что потребление корма в группах снижалось). Это может свидетельствовать о «зависании» в печени большого количества нелимитирующих аминокислот, которые оказываются невостребованными из-за отсутствия необходимого количества лизина и треонина. Такую же тенденцию, то есть значительное повышение суммарного содержания аминокислот в печени крыс при имбалансе лизина и триптофана в рационе, мы наблюдали ранее (23). В свою очередь, перегруженность печени аминокислотами и их метаболитами может быть причиной аноректической реакции. Это предположение в определенной мере подтверждают исследования зарубежных авторов по ваготомии. Так, субдиафрагматическая ваготомия, включая гепатическую и желудочную ветви блуждающего нерва, способствовала хорошему аппетиту у имбалансных крыс. Авторы полагают, что эта область вагусной иннервации может играть важную роль в начальной реакции животных на им-баланс (24). Возможно, первая информация об имбалансе аминокислот поступает в центральную нервную систему из печени и кишечника через блуждающий нерв, и на этой основе формируется соответствующее пищевое поведение животных.
Организм не накапливает аминокислоты, как жир или гликоген. Можно полагать, что возникающая при имбалансе аноректическая реакция, а также отсутствие или слабая адаптация к имбалансу обусловлены недостаточной способностью организма, в том числе печени, к деградации большого потока «лишних» аминокислот. Поэтому реакцию животных на имбаланс, которая проявляется в снижении аппетита или полном отказе от корма, следует рассматривать как физиологически обоснованный защитный рефлекс, обеспечивающий поддержание гомеостаза. То, что в мозге треонин-имбалансных цыплят на молекулярном уровне формируется система, вызывающая анорексию, вполне очевидно, поскольку в результате интраперитонеальной инъекции экстракта их мозга наблюдалось заметное снижение потребления корма у инъецированных особей. Нейропептид Y явно стимулировал потребление цыплятами треонин-имбалансного рациона. Поэтому проведение дальнейших исследований по молекулярной идентификации факторов регуляции аппетита при имбалансе представляет определенный интерес.
Таким образом, мы показали, что цыплята-бройлеры реагируют на им-баланс рациона по лизину и треонину снижением потребления корма и, как следствие, замедлением роста. Действие имбаланса определяется двумя факторами: дефицитом лимитирующей аминокислоты и «избытком» остальных аминокислот. При имбалансе лизина или треонина соответственно снижается их концентрация в плазме крови и содержание лизина в гипоталамусе, а содержание в печени и целом мозге остается таким же, как у цыплят на основном и скорректированном рационах. По-видимому, содержание аминокислот в мозге животных регулируется на гомеостатическом уровне. В печени у имбалансных цыплят заметно возрастает суммарное количество аминокислот, содержание мочевины и таурина, что свидетельствует о напряженных процессах по нейтрализации «лишних» аминокислот. Можно полагать, что образующиеся продукты деградации аминокислот (аммиак и мочевина) негативно действуют на обмен веществ и здоровье животных, вследствие чего снижение потребления пищи при имбалансе следует рассматривать как физиологически обоснованную защитную реакцию организма. Экстракты из целого мозга треонин-имбалансных цыплят, по-видимому, содержат вещества, способные вызывать аноректическую реакцию на соответствующий имбаланс. Идентификация этих факторов регуляции аппетита на молекулярном уровне представляет определенный интерес. Животные обладают чувствительным механизмом поиска и идентификации альтернативных источников дефицитных аминокислот. При смене местоположения этого источника цыплята быстро его находят и прочно удерживают в памяти. 56
Л И Т Е Р А Т У Р А
-
1. G i e t z e n D.W., E r e c i u s L.F., R o g e r s Q.R. Neurochemical changes after imbalanced diets suggest a brain circuit mediating anorectic responses to amino acid deficiency in rats. J. Nutr., 1998, 128: 771-781.
-
2. T r u o n g B.G., M a g n u m L.J., G i e t z e n D.W. GABA (A) and GABA (B) receptors in the
anterior piriform cortex modulate feeding in rats. Brain Res., 2002, 924 (1): 1-9.
-
3. K o e h n l e T.J., R u s s e l l M.C., G i e t z e n D.W. Rats rapidly reject diets deficient in essen
tial amino acid. J. Nutr., 2003, 133: 2331-2335.
-
4. K o e h n l e T.J., R u s s e l l M.C., M o r i n A.S. e.a. Diets deficient in indispensable aminoacids rapidly decrease the concentration of the limiting aminoacid in anterior piriform cortex of rats. J. Nutr., 2004, 134: 2365-2371.
-
5. E v e n P.C., R o l l a n d V., F e u r t e S. e.a. Postprandial metabolism and aversive response in rats fed threonine-devoid diet. Am. J. Physiol., 2000, 279: R248-R254.
-
6. Р я д ч и к о в В.Г. Улучшение зерновых белков и их оценка. М., 1978.
-
7. D a r c e l N., F r o m e n t i n G., R a y b o u l d H. e.a. Fos-positive neurons are increased in
the nucleus of the solitary tract and decreased in the ventromedial hypothalamus and аmygdala by a high-protein diet in the rats. J. Nutr., 2005, 135: 1486-1490.
-
8. H a r p e r A.E., B e n e v e n g a N.J., W o h l h u e t e r R.M. Effect of ingestion of disproportionate amounts of amino acids. Physiol. Rev., 1970, 50: 428-558.
-
9. G i e t z e n D.W., Ma g n u m L.J. Molecular mechanism in the brain involved in the anorexia of branched-chain amino acid deficiency. J. Nutr., 1999, 219: 1979-1983.
-
10. G i e t z e n D.W., R o s s C., H a o Sh. e.a. Phosphorilation of eIF2a is involved in the signaling of indispensable amino acid deficiency in the anterior piriform cortex of the brain in rats. J. Nutr., 2004, 134: 717-723.
-
11. H a o Sh., Sharp J., R o s s - I n t a C. e.a. Uncharged tRNA and amino acid deficiency in mammalian piriform cortex. Science, Mar., 2005, 307: 1776-1778.
-
12. C u m m i n g s S.L., T r u o n g B.G., G i e t z e n D.W. Neyropeptide Y and somatostatin in the
anterior piriform cortex after intake amino acid deficient diets. Peptides, 1988, 19: 527-535.
-
13. A n d o R., K a w a k a m i S.I., B u n g o T. e.a. Feeding responses to several neuropeptide Y
receptor agonist in the neonatal chick. Eur. J. Pharmacol., 2001, 427: 53-59.
-
14. T a c h i b a n a T.S., T a k a h a s h i H., U k e n a K. e.a. Gonadotropin-inhibiting hormone
stimulates feeding behavior in chicks. Brain Res., 2005, 1050(1-2): 94-100.
-
15. Рекомендации по кормлению птицы. Сергиев-Посад, ВНИТИП, 2003.
-
16. Р я д ч и к о в В.Г., Т а р а б р и н И.В., Р а д у л ь Н.П. и др. Пищевое поведение животных при разных формах баланса незаменимых аминокислот. С.-х. биол., 2005, № 2: 3-13.
-
17. Р я д ч и к о в В.Г. Обмен веществ у моногастричных животных при имбалансе аминокислот и пути повышения биологической ценности белка зерна злаковых культур. Докт. дис. Краснодар, 1981.
-
18. T e w s .K., G r e e n w o o d J., P r a t t O.E. e.a. Dietary aminoacid analogues and transport of lysine or valine across the blood-brain barrier in rats. J. Nutr., 1988, 118: 756-763.
-
19. C h u S.H.W., H e g s t e d D.M. Adaptive response of lysine and threonine degrading enzymes in adult rats. J. Nutr., 1976, 106: 1089-1096.
-
20. Y u a n J.-H., D a v i s A.J., A u s t i c R.E. Temporal response of hepatic threonine dehydrogenase in chickens to the initial consumption of a threonine imbalanced diet. J. Nutr., 2000, 130: 2746-2752.
-
21. T o r r e s N., B e r i s t a i n L., B o u r g e s H. e.a. Histidine-imbalanced diets stimulate hepatic histidase gene expression in rats. J. Nutr., 1999, 219: 1979-1983.
-
22. О м а р о в М.О. Биохимическое обоснование влияния некоторых незаменимых аминокислот в питании моногастричных животных на обмен веществ и продуктивность. Автореф. докт. дис. Краснодар, 2001.
-
23. Р я д ч и к о в В.Г., П л о т н и к о в В.К., П л о т н и к о в а А.В. Баланс аминокислот как регулятор аппетита и синтеза белка у свиней. В сб.: Повышение продуктивности свиноводства на Северном Кавказе. Краснодар, СКНИИЖ 1986: 39-57.
-
24. D i x o n K.D., W i l l i a m s F.E., W i g g i n s R.L. e.a. Differential effect of selective vagotomy and tropisetron in aminoprivic feeding. Am. J. Physiol. Regulative Comp. Physiol., 2000, 279: R997-R1009.
Кубанский государственный аграрныйуниверситет
