Реорганизация синаптоархитектоники сенсомоторной коры головного мозга у крыс линии Крушинского-Молодкиной после однократного шумового воздействия выше предельно допустимого уровня

Автор: Ерениев Степан Иванович, Семченко Валерий Васильевич, Степанов Сергей Степанович
Журнал: Вестник Омского государственного аграрного университета @vestnik-omgau
Рубрика: Ветеринарные науки
Статья в выпуске: 3 (23), 2016 года.
Бесплатный доступ
Установлено, что вследствие однократного шумового воздействия (ОШВ) выше предельно допустимого уровня в эксперименте на крысах линии К-М с использованием методов электронной микроскопии, морфометрического и статистического исследования происходит статистически значимое снижение общей численная плотности синапсов через 4, 8 и 24 ч соответственно на 67,7; 95,0 и 51,4 % от контроля. Кроме того, изменялось относительное содержание мелких, средних и крупных синапсов, синапсов с разной высотой плотных проекций, плоских, (+) и () искривленных контактов, изменялись форма и размеры шипиков, что свидетельствовало о существенной реорганизации синаптоархитектоники коры головного мозга экспериментальных животных после ОШВ. Все это может быть структурной основой психоэмоциональных и неврологических проявлений воздействия запредельного шума, а также основой формирования патологических систем мозга на базе гиперактивных нейронов и нейронных сетей мозга.
Шум выше предельно допустимого уровня, крысы линии крушинского - молод-киной, кора головного мозга, синаптархитектоника
Короткий адрес: https://sciup.org/142199237
IDR: 142199237 | УДК: 613.644:612.825-091.8-092.8
Synaptoarchitectonics reorganization of the sensorimotor cortex in rats of Krushinsky-Molodkina line after a single soundimpact exceeding maximum acceptable level
We studied the effects of a single sound exposure (SSE) maximum permissible level on synapses in layer I of the rats sensorimotor cortex (n = 90, Krushinsky-Molodkina line). Used methods of electron microscopy, morphometric and statistical analysis. After the SNE of synapses total numerical density was significantly reduced and after 4, 8 and 24 h, respectively 67.7; 95.0 and 51.4 % of the control. In addition, changed the relative content of small, medium and large synapses, synapses with different height of dense projections, flat, (+) and () curved contacts, changed the shape and size of spines. All this testified to a significant reorganization of the experimental animals cerebral cortex synaptoarchitectonics after the SSE. All this can be structural basis of psycho-biosocial and neurological manifestations of the impact of the noise exceeding the maximum value, as well as basis for the formation of pathological brain systems based on the hyperactive neurons and brain neural networks.
Текст научной статьи Реорганизация синаптоархитектоники сенсомоторной коры головного мозга у крыс линии Крушинского-Молодкиной после однократного шумового воздействия выше предельно допустимого уровня
В связи с тем, что в России заболевания, связанные с воздействием вредных физических факторов производства, наиболее распространенным из которых является шум [1, 2, 3], занимают наибольший удельный вес в структуре хронических профессиональных заболеваний и включены в «Перечень профессиональных заболеваний» [4]. Представляет интерес экспериментальное изучение воздействия шума, превышающего ПДУ, на синаптоархитектонику КГМ. Установлено, что шумовые воздействия вызывают нарушение долговременной памяти, способности к обучению и поведенческие реакции у крыс Вистар и линии Крушинского – Молодкиной [5].
Объекты и методы
В качестве экспериментальных животных использовали крыс линии Крушинского – Молодкиной (К-М; n = 90) с генетически детерминированной высокой чувствительностью к шумовому воздействию (ШВ), предоставленных виварием лаборатории физиологии и генетики
поведения кафедры высшей нервной деятельности биолого-почвенного факультета МГУ им. М.В. Ломоносова. Опыты проводили в соответствии с приказами МЗ СССР № 755 от 12.08.77 и № 701 от 27.07.78 об обеспечении принципов гуманного обращения с экспериментальными животными. Однократное шумовое воздействие (ОШВ) превышало предельно допустимый уровень (ПДУ = 80 дБ А) и равнялось 86 дБ А. Реакцию на шумовой раздражитель оценивали по методу Л.В. Крушинского [6] в звукоизолированной прозрачной камере (40 × 60 × 50 см), позволяющей вести визуальное наблюдение за двигательной активностью и психосоматическим состоянием животных.
Для электронно-микроскопического исследования кору головного мозга (КГМ) крыс (поле Fpa и Fpp) фиксировали погружением в смеси 1 %-ного раствора глютарового альдегида и 4 %-ного раствора параформа на 0,1 М фосфатном буфере (рН 7,4) с добавлением сахарозы (5 %). Затем материал отмывали в фосфатном буфере в течение 1 ч, дофиксировали в 1 %-ном растворе четырехокиси осмия в течение 2 ч, обезвоживали в спиртах восходящей концентрации и заключали в смесь эпона и аралдита. Тангенциальные ультратонкие срезы коры контрастировали уранилацетатом и цитратом свинца. Осмированный материал использовали для качественной и количественной оценки состояния нейронов, их отростков, синапсов и глиоцитов. Для селективного выявления синаптического цитоскелета ССЕ (система субсинаптических единиц) - плотные проекции, постсинаптическое уплотнение – использовали контрастирование в 1 %-ном растворе фосфорно-вольфрамовой кислоты (ФВК) на абсолютном спирте (100 %) (ФВК) [7, 8]. Фотографировали на электронном микроскопе 100 АКА. На электронограм-мах определяли общую численную плотность синапсов (ОЧПС) на 100 мкм2 нейропиля, выделяли симметричные, асимметричные и смешанные синапсы, плоские, положительно и отрицательно искривленные, неперфорированные и перфорированные, синапсы с низкими, средними и высокими плотными проекциями (типы А, В, С), очень мелкие, мелкие, средние, крупные и очень крупные. Вычисляли площадь активной зоны (АЗК), площадь свободной от АЗК поверхности шипика (СПШ), отношение СПШ/АЗК, средний диаметр АЗК, суммарную длину АЗК на 100 мкм2 нейропиля [7, 8]. Случайным образом выбранные участки фотографировали при стандартном увеличении в 15000 раз. Обработку материала проводили на цифровых изображениях негативов с помощью программы ImageJ 1.46. В качестве контроля использовали крыс линии К-М без ШВ.
Статистический анализ полученного материала проводили с использованием стандартного пакета прикладных статистических программ Statistica 6.0 [9]. Проверку статистических гипотез проводили с помощью t-критерия Стьюдента для независимых выборок. Нулевая гипотеза отвергалась при p < 0,05.
Результаты исследований и обсуждение
У животных контрольной группы среди функционально зрелых контактов, которые составляли 82,2 % от общего количества синапсов, содержание плоских, вогнутых и выпуклых контактов 1,4 : 1,1 : 1,0. Незрелые контакты были представлены только плоскими синапсами и составляли 17,8 % от общего количества (табл. 1). Численная плотность симметричных и асимметричных контактов 1 : 4,6. Преобладали функционально зрелые синапсы групп А и В мелких и средних размеров (табл. 2 и 3). ССЕ этих синапсов имела четкие контуры. Отношение СПШ/АЗК равнялось 2,55 (табл. 4). ССЕ имела структурную связь с окружающим ФВК-позитивным материалом цитоскелета пре- и постсинаптической зоны синапсов. После ОШВ ОЧПС уменьшалась и через 4, 8 и 24 ч составляла соответственно 67,7; 95,0 и 51,4 % от контроля. Редукция синапсов осуществлялась за счет всех разновидностей симметричных и асимметричных контактов (табл. 1).
Через 4 ч после ОШВ численная плотность синапсов снижалась на 32,3 %. На 23,2 % увеличивалось относительное содержание положительно искривленных контактов, а содержание плоских форм зрелых контактов снижалось на 13,6 % (рисунок). Относительное содержание асимметричных отрицательно искривленных и симметричных плоских контактов сохранялось на контрольном уровне. Уменьшалось содержание очень мелких, мелких синапсов и синапсов с высотой ПП 50 нм и менее, увеличивалось содержание очень крупных синапсов. Уменьша- лось отношение СПШ/АЗК, укорочение и утолщение шейки дендритов. Выявлялись синапсы с деструктивно измененными ПП: снижалась четкость контуров ПП, их высота, фрагментация и отрыв ПП от пресинаптической мембраны. В некоторых синапсах часть ПП контакта разрушалась полностью. Постсинаптическое уплотнение (ПСУ) повреждалось в меньшей степени.
Таблица 1
Численная плотность контактов с различной организацией системы субсинаптических единиц в нейропиле слоя I сенсомоторной коры головного мозга у крыс линии Крушинского – Молодкиной после однократного шумового воздействия, M ± m
|
Показатель |
Контроль |
Время после ОШВ, часы |
||
|
4 |
8 |
24 |
||
|
ОЧПС на 100 мкм2 нейропиля |
17,9 ± 0,41 |
11,97 ± 0,12* |
16,79 ± 0,30 |
9,10 ± 0,11* |
|
Асимметричные: всего плоские (+) искривленные ( - ) искривленные |
14,54 ± 0,28 5,88 ± 0,42 4,58 ± 0,23 4,08 ± 0,11 |
10,50 ± 0,15* 2,35 ± 0,12* 5,87 ± 0,21* 2,28 ± 0,13* |
15,61 ± 0,41 5,75 ± 0,33 5,87 ± 0,31 4,02 ± 0,58 |
8,05 ± 0,21* 3,49 ± 0,20* 2,38 ± 0,12* 2,28 ± 0,30* |
|
Симметричные: всего плоские |
3,15 ± 0,20 3,15 ± 0,20 |
1,41 ± 0,11* 1,41 ± 0,11* |
1,18 ± 0,11* 1,18 ± 0,11* |
1,05 ± 0,21* 1,05 ± 0,21* |
|
Диаметр АЗК, нм |
372,8 ± 34,3 |
434,7 ± 40,0 |
430,3 ± 39,6 |
584,4 ± 53,8* |
|
Суммарная длина АЗК, нм |
6595 ± 608 |
5203 ± 479 |
7225 ± 665 |
5318 ± 490 |
Таблица 2
Численная плотность синапсов с различной длиной активной зоны контакта в молекулярном слое сенсомоторной коры головного мозга у крыс линии Крушинского – Молодкиной в различные сроки после однократного шумового воздействия (ФВК-метод ), M ± m
|
Длина АЗК, нм |
Контроль |
Время после ОШВ, часы |
||
|
4 |
8 |
24 |
||
|
< 200 |
1,01 ± 0,02 |
0,46 ± 0,05* |
0,77 ± 0,01* |
0,29 ± 0,04* |
|
201 – 400 |
7,92 ± 0,52 |
3,56 ± 0,30* |
6,01 ± 0,01* |
0,47 ± 0,01* |
|
401 – 600 |
4,50 ± 0,23 |
3,81 ± 0,40 |
3,42 ± 0,06* |
1,54 ± 0,02* |
|
601 – 800 |
3,24 ± 0,39 |
2,73 ± 0,30 |
4,61 ± 0,08* |
4,17 ± 0,05* |
|
> 800 |
1,02 ± 0,04 |
1,41 ± 0,01* |
1,98 ± 0,04* |
2,63 ± 0,03* |
Таблица 3
Количество асимметричных контактов с различной высотой плотных проекций в слое I сенсомоторной коры головного мозга у крыс линии Крушинского – Молодкиной в различные сроки после однократного шумового воздействия (ФВК-метод), M ± m
|
Время после ОШВ, часы |
Количество синапсов с различной высотой плотных проекций на 100 мкм2 нейропиля |
||
|
> 60 нм |
60–51 нм |
50 нм и < |
|
|
Контроль |
6,26 ± 0,37 |
5,04 ± 0,29 |
3,24 + 0,16 |
|
4 |
3,01 + 0,18* |
5,46 + 0,31 |
2,03 ± 0,10* |
|
8 |
5,04 ± 0,30 |
7,21 + 0,41 * |
3,36 + 0,17 |
|
24 |
3,92 ± 0,34* |
2,09 + 0,15* |
2,04 + 0,14* |
Таблица 4
Дендритные шипики слоя I сенсомоторной коры головного мозга у крыс линии Крушинского – Молодкиной в различные сроки после однократного шумового воздействия, M ± m
|
Показатель |
Контроль |
Время после ОШВ, часы |
||
|
4 |
8 |
24 |
||
|
Площадь АЗК, нм2 |
109111 ± 926 |
148320 ± 1259* |
145359 ± 1233* |
268091 ± 2275* |
|
СПШ, нм2 |
278233 ± 2361 |
344102 ± 2919* |
382295 ± 3243* |
726527 ± 6164* |
|
СПШ/АЗК |
2,55 |
2,32 |
2,63 |
2,72 |
Примечание. В табл. 1 – 4 символ «*» обозначает, что в сравнении с контролем различия статистически значимы при р < 0,05 (t – критерий Стьюдента для независимых выборок). АЗК – активная зона контакта; ОШВ – однократное шумовое воздействие; ПП – плотные проекции; СПШ – свободная поверхность шипика; ОЧПС – общая численная плотность синапсов. Материал представлен как среднее (M) ± стандартная ошибка средней (m).
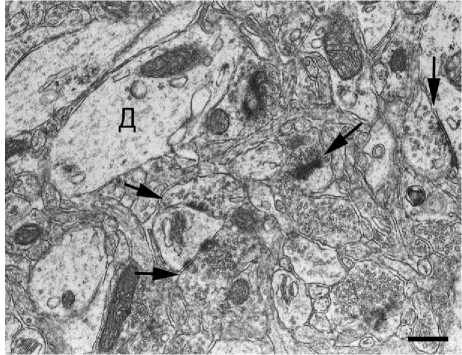
А
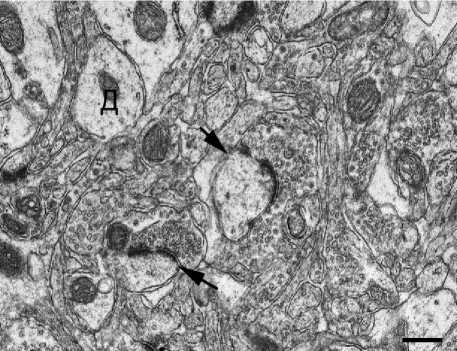
Б
Нейропиль сенсомоторной коры головного мозга у крыс линии Крушинского – Молодкиной:
А - контроль, преобладают плоские контакты; Б - через 4 часа после однократного шумового воздействия выше предельно допустимого уровня; увеличение относительного содержания положительно искривленных функционально зрелых синапсов; Д - дендрит; стрелки - синапсы. Контрастирование цитратом свинца и уранилацетатом после дофиксации 1 %-ным раствором четырехокиси осмия. Ув. × 18000, шкала - 500 нм
Через 8 ч после ОШВ общее количество выявляемых межнейронных контактов на 100 мкм2 снижалось на 5,1 %. Относительное содержание положительно искривленных контактов уменьшалось на 14,1 % по сравнению с предыдущим сроком, а содержание плоских контактов возрастало на 14,6 %. При этом по сравнению с контролем на 10,8 % уменьшалось содержание плоских симметричных контактов, а содержание отрицательно искривленных оставалось на уровне контроля. Увеличивалось количество крупных и очень крупных контактов, уменьшалось содержание синапсов с очень мелкой, мелкой и средней АЗК, увеличивались число синапсов с высотой ПП 51–60 нм, площадь АЗК и свободной от АЗК поверхности шипика, удлинялась и истончалась шейка дендритных шипиков.
Через 24 ч после ОШВ численная плотность выявляемых межнейронных контактов снижалась на 48,6 %. Относительное содержание положительно искривленных контактов становилось равным контролю, а количество плоских синапсов не отличалось от предыдущего срока и контрольного уровня. К концу суток после пароксизма происходило восстановление относительного содержания функционально зрелых и незрелых контактов, хотя абсолютное количество выявляемых синапсов на 100 мкм2 оставалось ниже, чем в контроле. Увеличивался средний диаметр контактов за счет увеличения количества крупных и очень крупных синапсов. Как и в контроле преобладали синапсы с высотой ПП более 60 нм. Возрастала площадь АЗК и свободной от АЗК поверхности шипика, удлинялась и истончалась шейка дендритных шипиков.
Закл ючение
Таким образом, ОШВ выше предельно допустимого уровня сопровождается изменениями синаптоархитектоники сенсомоторной КГМ, которые к концу суток после ОШВ не проходят полностью. Уменьшается численная плотность контактов. В первые часы после ОШВ часть межнейронных контактов из функционально неактивного состояния переходят в активное состояние передачи нервного импульса, что документируется увеличением относительного содержания положительно искривленных и уменьшением содержания плоских функционально зрелых синапсов. Активированное состояние межнейронных связей сохраняется в течение 4 ч после ОШВ и относительно нормализуется к концу суток. Через 24 ч после ОШВ увеличивается средний диаметр сохранившихся синапсов, уменьшается суммарная длина контактов на 100 мкм2 нейропиля, увеличивается относительное содержание крупных и очень крупных синапсов и синапсов с высотой ПП 51–60 нм. Восстановление синаптического пула происходит посредством трансформации незрелых плоских симметричных контактов в функционально зрелые асимметричные плоские контакты.
Выводы
Однократное шумовое воздействие, превышающее предельно допустимый уровень, вызывает качественные и количественные изменения синапсов в слое I сенсомоторной коры головного мозга у крыс линии Крушинского – Молодкиной. Через 4 часа после воздействия увеличивается содержание активных (положительно искривленных) синаптических контактов. Через 24 ч увеличивается средний диаметр сохранившихся синапсов, относительное содержание крупных и очень крупных синапсов и синапсов с высотой плотных проекций 51–60 нм, уменьшается суммарная длина контактов на 100 мкм2 нейропиля. Частичное восстановление синаптического пула к концу суток после шумового воздействия происходит за счет превращения незрелых плоских симметричных контактов в функционально зрелые асимметричные контакты.
Список литературы Реорганизация синаптоархитектоники сенсомоторной коры головного мозга у крыс линии Крушинского-Молодкиной после однократного шумового воздействия выше предельно допустимого уровня
- Профессиональная патология: национальное руководство/под ред. Н.Ф. Измерова. М.: ГЭОТАР-Медиа, 2011. 784 с.
- Профессиональные заболевания/под ред. Н.Ф. Измерова. М.: Медицина, 1996. Т. 2. С. 162-175.
- Российская энциклопедия по медицине труда/под ред. Н.Ф. Измерова. М.: Медицина, 2005. 656 с.
- Приказ МЗ и СР РФ от 27 апреля 2011 г. № 417н «Об утверждении перечня профессиональных заболеваний», приложение 1, раздел 2, пункты 2.4.1 и 2.4.2.
- Долговременная память и способность к обучению крыс с высокой судорожной активностью мозга при аллотрансплантации эмбриональной нервной ткани/С.И. Ерениев, В.В. Семченко, Р.И. Генне, К.К. Маковецкий//Журн. высшей нервн. деят. 1993. Т. 43, вып. 5. С. 987-993.
- Крушинский Л.В. Формирование поведения животных в норме и патологии. М.: Изд-во МГУ, 1960. 264 с.
- Synaptic remodelling during development and maturation: junction differentiation and splitting as a mechanism for modiffing connectivity/S.E. Dyson, D.G. Jones//Dev Brain Res. 1984. V. 13, № 1. P. 125-137.
- Семченко В.В., Степанов С.С., Боголепов Н.Н. Синаптическая пластичность головного мозга (фундаментальные и прикладные аспекты). М., 2014. 2-е изд. 499 с.
- Реброва О.Ю. Статистический анализ медицинских данных. Применение пакета прикладных программ STATISTICA. М.: МедиаСфера, 2002. 312 с.
