Скрининг гаплотипа фертильности АН1 айрширской породы крупного рогатого скота центрального и северо-западного регионов России

Автор: Гладырь Елена Александровна, Терновская Ольга Андреевна, Костюнина Ольга Васильевна
Журнал: АгроЗооТехника @azt-journal
Рубрика: Разведение, селекция и генетика сельскохозяйственных животных
Статья в выпуске: 4 т.1, 2018 года.
Бесплатный доступ
Айрширская порода является высокоспециализированной породой молочного скота, отличается высокой молочной продуктивностью, устойчивостью к инфекционным заболеваниям, продуктивным долголетием, хорошими показателями фитнеса и воспроизводства, конверсией корма и обладает пластичностью генофонда. Современный генофонд айрширского скота может содержать кровь пород красного корня: красной шведской, красной норвежской, айрширской американского происхождения, часто получаемых с прилитием крови красно-пестрой голштинской породы, а также швицкой породы. В породе наблюдается тенденция снижения фертильности коров и выхода телят, вызванная LOF-мутациями, приводящими к эмбриональным потерям или постнатально проявляющимися как генетические дефекты. PIRM синдром айрширского скота, проявляющийся в нарушении развития и ранней смерти молодняка в айрширских стадах, обусловлен гомозиготным состоянием гена UBE3B по мутантному аллелю, является аутосомальным, рецессивными дефектом, приводящим к появлению гаплотипа фертильности АН1 айрширского скота. Изучение распространенности гаплотипа фертильности АН1 в российской популяции айрширского скота Центрального и Северо-Западного регионов России, составлявшее цель работы, было осуществлено посредством решения ряда задач (теоретическое моделирование и практическая отработка аллелеспецифической полимеразной цепной реакции (АС-ПЦР), экспериментальное тестирование животных айрширской породы и последующий мониторинг состояния распространения гаплотипа АН1 в популяции айрширского скота четырех областей России). Исследование 259 коров и 28 быков-производителей, разводимых в четырех областях Центрального и Северо-Западного регионов России, выявило наличие 47 скрытых носителей АH1, в том числе 37 коров и 10 быков, что соответствует частотам встречаемости, 14,3 и 35,7%, соответственно, и в среднем по породе - 16,4%. Быки-производители пяти племпредприятий, носители АН1 гаплотипа, принадлежат в основном линии О.Р. Лихтинг (Лайтнинг) 120135, код 19. Результаты исследования полиморфизма гена UBE3B и частоты встречаемости носителей АН1 гаплотипа в племенных стадах айрширского молочного скота России найдут свое применение в селекционно-племенной работе с породой.
Айрширская порода, lof-мутации, ген ube3b, гаплотип фертильности ан1
Короткий адрес: https://sciup.org/147225576
IDR: 147225576 | УДК: 636.2.082.2 | DOI: 10.15838/alt.2018.1.4.1
Screening of АH1 fertility haplotype in the Ayrshire cattle breed in the central and northwestern regions of Russia
The Ayrshire breed is a highly specialized breed of dairy cattle, characterized by high milk production, resistance to infectious diseases, productive longevity, good fitness, reproduction and feed conversion and has plasticity of the gene pool. The modern gene pool of the Ayrshire cattle may contain the blood of red breeds: Swedish Red, Norwegian Red, Ayrshire of American origin, often derived from admixture of the blood of the Red-and-White Holstein and Brown Swiss breeds. In the breed, there is a tendency toward decreasing the fertility of cows and calves, caused by LOF-mutations that lead to embryonic losses or that are manifested postnatally as genetic defects. PIRM syndrome of the Ayrshire cattle, manifested in the violation of development and early death of young animals in Ayrshire herds, is due to the homozygous state of the gene UBE3B for the mutant allele and is an autosomal and recessive defect leading to the appearance of AH1 fertility haplotype in the Ayrshire cattle. The goal of our work was to study the prevalence of AH1 fertility haplotype in the Russian population of the Ayrshire cattle in theCentral and Northwestern regions of Russia; the work was carried out by solving a number of problems (theoretical modeling and practical testing of allele-specific polymerase chain reaction, experimental testing of animals of the Ayrshire breed, and subsequent monitoring of the distribution of AH1 haplotype in the population of the Ayrshire cattle in four regions of Russia). The study of 259 cows and 28 seed bulls bred in four regions of the Central and Northwestern regions of Russia revealed the presence of 47 hidden carriers of AH1, including 37 cows and 10 bulls, which corresponds to the frequency of occurrence equal to 14.3 and 35.7%, respectively, and on average in the breed - 16.4%. Seed bulls of five stud farms, carriers of AH1 haplotype, belong generally to the line of O.R. Lichting (Lightning) 120135, code 19. The results of the study of UBE3B gene polymorphism and the frequency of occurrence of AH1 haplotype carriers in breeding herds of the Ayrshire dairy cattle in Russia will be used in breeding work with the breed.
Текст научной статьи Скрининг гаплотипа фертильности АН1 айрширской породы крупного рогатого скота центрального и северо-западного регионов России
АН1 айрширского скота. Изучение распространенности гаплотипа фертильности АН1 в российской популяции айрширского скота Центрального и Северо-Западного регионов России, составлявшее цель работы, было осуществлено посредством решения ряда задач (теоретическое моделирование и практическая отработка аллелеспецифической полимеразной цепной реакции (АС-ПЦР), экспериментальное тестирование животных айрширской породы и последующий мониторинг состояния распространения гаплотипа АН1 в популяции айрширского скота четырех областей России). Исследование 259 коров и 28 быков-производителей, разводимых в четырех областях Центрального и Северо-Западного регионов России, выявило наличие 47 скрытых носителей АH1, в том числе 37 коров и 10 быков, что соответствует частотам встречаемости, 14,3 и 35,7%, соответственно, и в среднем по породе – 16,4%. Быки-производители пяти племпредприя-тий, носители АН1 гаплотипа, принадлежат в основном линии О.Р. Лихтинг (Лайтнинг) 120135, код 19. Результаты исследования полиморфизма гена UBE3B и частоты встречаемости носителей АН1 гаплотипа в племенных стадах айрширского молочного скота России найдут свое применение в селекционно-племенной работе с породой.
Айрширская порода, LOF-мутации, ген UBE3B, гаплотип фертильности АН1.
Айрширская порода является высокоспециализированной породой молочного скота, разводится в ряде стран мира и занимает свое отдельное место среди молочных пород Старого и Нового Света. Порода отличается высокой молочной продуктивностью и качеством молока, резистентностью к различным инфекционным заболеваниям, продуктивным долголетием, прекрасными адаптационными качествами, показателями фитнеса и воспроизводства, лучшей конверсией корма и обладает пластичностью генофонда [1, с. 1; 2, с. 22]. Суммарное поголовье животных айрширской породы в России образует вторую после Финляндии по размерам мировую популяцию айршир-ского скота и насчитывает 77,67 тыс. голов, в том числе 50,82 тыс. гол. коров, что составляет 3,05% от всех коров молочных и молочно-мясных пород страны [3, с. 15]. В базе данных ВНИИПлем по национальному генофонду сельскохозяйственных животных – племенного крупного рогатого скота молочного направления продуктивности в 2018 году содержится информация о 9099 коровах [4], разводимых в 12 регионах страны, и 1017 быках [5].
Пожизненная молочная продуктивность животных айрширской породы, разводимых в племенных заводах России, сформированная с учетом экономической оценки по сумме количества молочного жира и белка за весь период использования животного, позволяет породе войти в пятерку лидирующих молочных пород России [6, с. 17; 7, с. 101–102].
В современной селекции айрширского скота России наряду с применением метода чистопородного разведения, позволившего получить заводской тип «Новоладожский», животные которого отличаются более высокой продуктивностью, живой массой, приспособленностью к промышленной технологии и устойчивостью к лейкозу [8, с. 5], используют кроссбред-ных быков пород красного корня: красной шведской, красной норвежской [9, с. 188], айрширской американского происхождения, часто получаемых с прилитием крови красно-пестрой голштинской породы [10, с. 2; 11, с. 1]. На долю неродственных пород, таких как бурая швицкая, голштинская и фризская, приходится менее 1% [12, с. 17]. Ранее было показано, что в айрширских стадах Швеции наряду с традиционными финскими линиями существуют родственные группы быков норвежского происхождения и используются производители разного направления продуктивности, что приводит к существенным фенотипическим отличиям животных, при этом некоторые производители несут кровь быков швицкой породы [13, с. 11–21].
При сохраняющемся чистопородном разведении и ведении племенных книг по породам в селекционных программах Европы все чаще используется кроссбридинг, при котором финская айрширская, красная датская и красная шведская начиная с 2010 года объединяются как совокупная композитная порода, имеющая во многом схожую селекционную историю, и разводятся в единой селекционно-племенной программе, называемой красная нордическая (Viking Red), геномно оцениваются вместе, получая единую генетическую оценку [14, с. 4486; 15, с. 13–14]. Красная норвежская порода генетически и селекционно связана с красной шведской и айрширской финской селекции [16, с. 472], в то время как красную датскую, в которой присутствуют гены не менее 8 пород, в том числе монбельярдской – 2,7%, голштинской – до 19,8% [12, с. 18], финскую айрширскую и красную норвежскую используют в селекционных программах разведения красной шведской породы [17, c. 3]. Вследствие этого быки, получаемые в Северной Европе посредством обмена генетического материала между популяциями красных скандинавских пород, приобретают генетическое родство между собой, и совместная обработка генетических данных в единой базе способствует повышению точности их геномных оценок [14, с. 4485], что ранее было показано для других пород крупного рогатого скота [18, c. 8, 19, с. 75]. Кроме того, ряд стран, такие как США, Голландия, Италия, Германия, Португалия и Австралия, работает в системе ПроКросс, используя трехпородное ротационное скрещивание голштинской, красной нордической и монтбельярдской пород [20, с. 1; 21, с. 1–3]. Эксперименты по изучению фертильности, устойчивости к маститам и молочной продуктивности кроссбредных животных красных скандинавских пород с голштинской в сравнении с чистопородными голштинскими коровами в течение пяти лактаций показали существенное улучшение показателей фертильности, здоровья и совокупной молочной продуктивности в группе кроссбредных животных [22, с. 918].
Вместе с тем в породе наблюдается тенденция снижения фертильности коров и выхода телят, вызванная генетическими причинами [23, c. 1; 24 с. 895; 25, с. 1; 26, с. 1], что является актуальной проблемой не только айрширского скота. Ранее показано, что снижение репродуктивных показателей, обусловленное генетическими факторами, названными гаплотипами фертильности, играет все более значимую роль в современном молочном скотоводстве в селекции основных молочных пород скота: голштинской и родственной ей черно-пестрой [27, с. 1; 28, с. 30; 29, с. 430–431; 30, с. 82–83], бурой швицкой [23, с. 1; 31, с. 6153; 32, с. 400], джерсей-ской [33, с. 4; 34, с. 1], симментальской [35, с. 1], монтбельярдской [36, с. 5], красной норвежской [37, с. 7–8; 38, с. 19–23]. Так, в геноме человека из аннотированных генов 7168 (33,3%) имеют важное значение и функциональность по меньшей мере одной копии каждого из этих генов жизненно необходима [39, с. D940]. Большинство LOF-мутаций приводит к ранним эмбриональным потерям из-за невозможности имплантации и дальнейшего развития эмбриона, но есть и такие, которые проявляются во второй и третий триместр беременности или постнатально как генетические дефекты [40, c. 1–2]. Экономические потери от LOF-мутаций в США оценены в 10,74 млн дол. и вызваны известными рецессивными летальными мутациями в основных четырех молочных породах скота: голштинской, джерсейской, бурой швицкой и айрширской, причем на долю первой из них приходится примерно 70% всех убытков [41, с. 7274]. Следовательно, понимание геномной архитектуры популяций крупного рогатого скота становится все более важным теперь, чем когда-либо, ввиду сложных комбинативных сочетаний генотипа животных для прогнозирования генетического прогресса, сдерживания отрицательного воздействия вредных мутаций, ответственных за генетические дефекты и инбредную депрессию [42, с. 49–58]. Возможности полногеномных исследований (WGS) в выявлении не только единичных замен (SNP) или небольших инсерций и делеций типа индел, но и фрагментов ДНК длиной более 50 п.о., представленных делециями, инсерциями, дупликациями, инверсиями и транслокациями, помогают исследованию наименее изученных структурных вариантов полиморфизмов крупного рогатого скота, которые, как считают, в значительной степени способствуют фенотипической вариабельности и имеют широкий диапазон воздействий от полезного до летального у разных видов сельскохозяйственных животных [43, с. 1–2]. Фенотипическим проявлением структурных вариантов можно объяснить дополнительную генетическую дисперсию, возникшую с изменением числа копий генов и ассоциированную с выходом молока, количества жира и белка [44, с. 1–8], и в то же время проявлением летального эффекта больших делеций у молочного скота разных пород [37, с. 6–8; 45, с. 1; 46, с. 1–2; 47, с. 10–11]. Применение в системе воспроизводства ограниченного числа высокоценных быков сопряжено с риском быстрого распространения рецессивных генетических дефектов в стадах породы [28, с. 30–31].
Именно летальным эффектом проявляется гомозиготная мутация в гене UBE3B крупного рогатого скота, расположенная на 17 BTA в диапазоне 65,9–66,2 М п.о. (сборка генома UMD 3.1 assembly) и приводящая к возникновению гаплотипа АН1 айрширского скота [48, с. 3881]. PIRM синдром айрширского скота, проявляющийся в нарушении развития и ранней смерти молодняка в айрширских стадах, представляет следствие гомозиготного состояния гена UBE3B по мутантному аллелю, является аутосомальным, рецессивным дефектом, возникновение которого обусловлено заменой последнего нуклеотида G > A в экзоне 23 гена UBE3B (rs475678587), приводящего к синонимичной замене E692E [24, с. 3, 49]. Показано, что изменение последовательности гена UBE3B приводит к нарушению сплайсинга и синтезу измененного белка убиквитин протеин лигаза E3B, обусловливающим способность специфически связываться с предназначенными для деградации белковыми субстратами и запускать сам процесс деградации, что приводит к нарушению клеточного гомеостаза [24, c. 2]. Анализ транскрипции UBE3B выявил пропуски экзонов у пораженных животных, что влекло за собой изменение белка, потерю в нем 40 аминокислот, из которых 20 находятся в сохраненном HECT-домене, каталитическом сайте белка UBE3B [50, с. 1]. Изучение родословных выявленных скрытых носителей АH1 показало, что вероятный самый старый носитель мутации – выдающийся бык-производитель USA117936 Selwood Betty’s Commander (1953 г. р.), являющийся наиболее часто используемым быком в породе, при этом ожидаемый уровень инбридинга по нему составил 11,1% [51, с. 2771, 2776]. T.A. Cooper с соавт. [48, с. 3878] было показано, что процент оплодотворения (SCR) в 618 спариваниях быков носителей АН1 с коровами, чьи отцы были носителями, снижается на 4,3±2,5%. Одним из эффектов летального действия гаплотипа АН1 является рост числа мерт- ворожденных телят на 1,4 и 2,1%, соответственно, от спариваний быков-носителей с коровами и телками, чьи отцы были носителями АН1. Вместе с тем значительного отрицательного эффекта от АН1 гаплотипа относительно ранней эмбриональной смертности плода (NRR) не наблюдалось [52, с. 102, 109]. Частота встречаемости носителей АН1 остается стабильно высокой с середины 70-х годов ХХ века, составляет примерно 26,1% в течение большого числа поколений и является самым распространенным рецессивным гаплотипом в мире [48, с. 3881; 50, с. 1].
Цель работы состояла в изучении распространенности гаплотипа фертильности АН1 в российской популяции айрширско-го скота Центрального и Северо-Западного регионов России с использованием созданной тест-системы. Задачами исследований было предусмотрено теоретическое моделирование и практическая апробация тест-системы на основе АС-ПЦР, экспериментальное тестирование животных айрширской породы и последующий мониторинг состояния распространения гаплотипа АН1 в популяции айрширского скота четырех областей России.
Новизна исследования заключается в том, что впервые была предпринята попытка оценки масштаба распространения гаплотипа АН1 четырех региональных популяций айрширского скота страны с использованием оригинальной АС-ПЦР.
Практическая значимость полученных результатов состоит в интеграции знаний о скрытых носителях АН1 в региональные программы разведения быков-производителей и коров айрширской породы.
Материалы и методы
Материалом для исследований служили пробы ткани (ушной выщип, кровь, сперма) от 287 предварительно здоровых особей (коров и быков) айрширской породы семи племенных предприятий РФ, архи- вированные в 2008–2017 годах. Выделение ДНК проводили с помощью наборов «ДНК-Экстран» (ЗАО «Синтол», Россия) и COrDIS (ООО «ГОРДИЗ», Россия) согласно стандартным протоколам производителей с некоторыми нашими модификациями. Подбор праймеров осуществляли с использованием программного обеспечения Primer3web, v. 4.0.0 [53, с. 1] с учетом следующих условий: минимальная, желаемая и максимальная температура отжига – +56, +60 и +64°С, соответственно; минимальное, желаемое и максимальное соотношение GC – 50, 60 и 70%, соответственно; длина фрагмента – 100–350 п.о. Постановку ПЦР осуществляли по методике Н.А. Зиновьевой с соавт. [54, с. 15]. Амплификацию проводили в конечном объеме 15 мкл реакционной смеси, содержащей 1*ПЦР буфер (16,6 мМ(NH4)2SO4; 67,7 мМ Трис – HCl, pH = 8,8; 0,1 (v/v)Tween 20), 1,5 мМMgCl2, 200мкМ диоксинуклеозидтрифосфатов, 12–15 пмоль каждого из праймеров, 1 Ед Taq-полимеразы и 1 мкл тестируемой ДНК (20–50 пмоль/мкл). Продукты АС-ПЦР разделяли посредством электрофореза в 3% агарозном геле с добавлением бромида димидия и визуализировали под ультрафиолетовым светом. Для идентификации длин фрагментов использовали молекулярный маркер длины 100 п.о. (500х2) (ООО «Биосан», Россия).
Результаты исследования
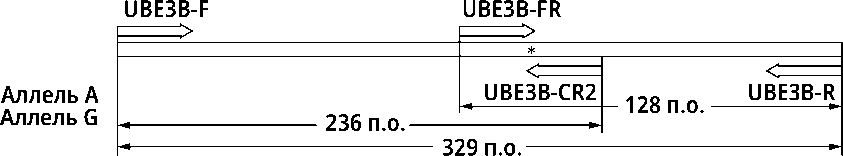
Определение полиморфизма гена UBE3B выполняли посредством однопробирочной АС-ПЦР с четырьмя праймерами, в которой праймеры UBE3B-F и UBE3B-R фланкировали общий для двух аллелей фрагмент длиной 329 п.о., а два «внутренних» аллелеспецифических праймера UBE3B-FR и UBE3B-CR2, ориентированные в противоположных направлениях, ам-плифицировали фрагменты, соответствующие аллелям G (236 п.о.) и A (128 п.о.) (рис. А).
A.
12 3 4 56789M
329 п.о. , 236 п.о 128п.о.
Б.
500 п.о.
400 п.о.
300 п.о.
200 п.о.
100п.о.
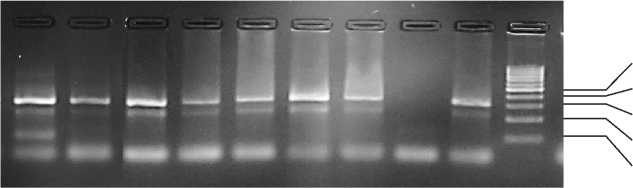
Рис. Теоретическая модель тест-системы определения полиморфизма UBE3B на основе АС-ПЦР (А) и результаты генотипирования образцов (Б)
А: UBE3B-F, UBE3B-R – праймеры, фланкирующие фрагмент; UBE3B-FR и UBE3B-CR2 – внутренние аллелеспецифические праймеры.
Б: дорожка 1 – генотип GA (скрытый носитель AH1); дорожки 2–7, 9 – генотип GG (не носитель AH1); дорожка 8 – отрицательный контроль амплификации (вода); М – маркер (длины «ступеней» маркера в парах оснований (п.о.) указаны справа от фотографии).
* Место исследуемой мутации.
Источник: экспериментальные расчеты и результаты авторов.
Анализ амплифицированных фрагментов в 3% агарозном геле, представленный на электрофореграмме на рис. Б, показывает, что генотипу GG по UBE3B соответствует наличие двух фрагментов длиной 329 и 236 п.о., в то время как генотипу GA – трех фрагментов длиной 329, 236 и 128 п.о. С применением созданной тест-системы исследовано 287 голов крупного рогатого скота – коров и быков айрширской породы, разводимых в четырех регионах Центральной и Северо-Западной России, при этом 240 образцов определено с генотипом GG (не носитель, АН1F) и 47 образцов с генотипом GA (скрытый носитель, AH1C). Генотип АА является летальным и не может быть выявлен среди взрослых животных.
Исследование 259 коров и 28 быков-производителей с использованием разработанной системы выявило наличие 47 скрытых носителей АH1, в том числе 37 коров и 10 быков, что соответствует частотам встречаемости носителей, соответственно,
14,3 и 35,7%, и в среднем по породе составило 16,4%. Быки-производители пяти племпредприятий, носители АН1 гаплотипа, принадлежат в основном линии О.Р. Лихтинг 120135. Ранее было сообщено, что наиболее часто носителями АН1 гаплотипа являются потомки канадских быков Jelyca Oblique, Des Chamois Poker – ET, DEs FLeurs Perfect – ET, De La Plaine Prime, ST Clement Edmour, Kildare Jupiter – ET, Kamouraska Bigstar – ET, Palmyra Bingo – ET, Duo Star Pokerstars, Des Coteaux Warner [55, с. 1; 56].
Полученные нами экспериментальные данные согласуются с результатами H. Venhoranta с соавт. [24, с. 3] по частоте встречаемости носителей АН1 гаплотипа в популяции финских айрширов (17,1%) и занимают промежуточное положение между данными Н.В. Ковалюк с соавт. [57, с. 16] и Null с соавторами [25, с. 1], 10,0 и 22,2%, в популяциях айрширских животных Краснодарского края и США, соответственно. В связи с достаточно высокой частотой встречаемости AH1 в исследованной нами выборке предложенная нами тест-система может быть рекомендована для массового скрининга племенного скота айршир-ской породы в РФ с целью обязательного мониторинга распространения скрытых носителей AH1. В заключение следует отметить, что современные селекционные программы, предусматривающие комби-нативное использование нескольких пород в создании промышленных кроссов, должны не только ориентироваться на эффект гетерозиса в получении желаемого селекционного результата за счет ком- бинации различных генотипов и генетических особенностей исходных пород, но и обращать пристальное внимание на контроль за распространением генетических дефектов, которыми обременены породы, для предотвращения трассирования летальных мутаций в будущие поколения.
Исследования проведены по теме государственного задания, регистрационный номер № АААА-А18-118021590138-1. В исследовании использовано оборудование ЦКП «Биоресурсы и биоинженерия сельскохозяйственных животных» ФГБНУ ФНЦ ВИЖ им. Л.К. Эрнста.
Список литературы Скрининг гаплотипа фертильности АН1 айрширской породы крупного рогатого скота центрального и северо-западного регионов России
- FABA. 2018. Available at: http://www.faba.fi/ru/dairy/breeding (accessed 30.09.2018).
- Генетический потенциал айрширского скота племенного завода «Новоладожский» Ленинградской области / О.В. Тулинова [и др.]. // Молочное и мясное скотоводство. 2015. № 5. С. 22-25.
- Ежегодник по племенной работе в молочном скотоводстве в хозяйствах Российской Федерации (2017 год) / И.М. Дунин [и др.]. М.: Изд-во ФГБНУ ВНИИплем, 2018. 274 с.
- ВНИИПлем. База данных национального генофонда. База данных по национальному генофонду сельскохозяйственных животных - племенного КРС молочного направления продуктивности. 2018. URL: http://xn-b1agiaphcs.xn-p1ai/grpzh (дата обращения 30.09.2018).
- ВНИИПлем. База данных быков по породам. Айрширская порода. 2017. URL: http://xn-b1agiaphcs.xn-p1ai/rus/files/Database/Bulls/2017/ajrshirskaya-poroda-bb.pdf (дата обращения 30.09.2018).
