Скрининг линий и сортов подсолнечника селекции ВНИИМК по устойчивости к ALS-ингибирующим гербицидам

Автор: Демурин Я.Н., Тронин А.С., Пихтярва А.А., Левуцкая А.Н., Фролов С.С., Гучетль С.З.
Рубрика: Селекция и семеноводство сельскохозяйственных растений
Статья в выпуске: 2 (159-160), 2014 года.
Бесплатный доступ
В задачу работы входило изучение частоты встречаемости генов гербицидоустойчивости в генофонде культурного подсолнечника ВНИИМК. Проведён широкомасштабный скрининг генофонда сортов и линий подсолнечника по устойчивости к гербицидам Пульсар и Экспресс. Исследования проводили на центральной экспериментальной базе ВНИИМК (г. Краснодар) и Армавирской опытной станции ВНИИМК (г. Армавир). Использовали сорта-популяции Родник, Мастер и Орешек, материнские линии ВК276, ВК678, ВА6 и ВА769, а также отцовские линии ВК580, ВК585 и ВА384. На трёх экспериментальных полях общей площадью 15 га при осмотре около 680000 растений трёх сортов и семи линий не было обнаружено ни одного типичного растения посеянных генотипов подсолнечника с признаком устойчивости к ALS-ингибирующим гербицидам: имидазолинонам или сульфонилмочевинам. Для молекулярной идентификации генов гебицидоустойчивости использовали ДНК-маркеры. ДНК подсолнечника выделяли из листьев индивидуальных вегетирующих растений по модифицированному методу Saghai-Maroof с соавторами. Оценивали электрофоретические спектры фрагментов ДНК подсолнечника, амплифициро- ванные с аллель-специфичными праймерами по гену ImiSun, а также с аллель-специфичными праймерами по гену CLHA-Plus. Частота встречаемости доминантного гена имидазолиноно- устойчивости оценивается менее чем 5 х 10 -6 (1 : 200000) для генофонда изученных сортов и линий ВНИИМК и менее 4 х 10 -6 для линий Армавирской опытной станции (1 : 280000). Частота встречаемости гена устойчивости к трибенурон- метилу у селекционных линий ВНИИМК не превышает значения 5 х 10 -6 (1 : 200000). Низкая частота встречаемости доминантных генов гербицидоустойчивости предполагает использование методов индуцированного мутагенеза для расширения наследственной изменчивости признака при поиске новых генов.
Резистентность, гербицид, имидазолиноны, сульфонилмочевины, линия, сорт
Короткий адрес: https://sciup.org/142151199
IDR: 142151199 | УДК: 633.854.78:575
Screening of sunflower lines and op varieties of VNIIMK breeding for ALS-inhibitor resistance
The main task of the research was to estimate the frequency of a dominant gene for resistance against herbicides in the oil bearing germplasm of VNIIMK. Wide screening for Pulsar and Express resistance of sunflower lines and open pollinated varieties of VNIIMK breeding was done. The research has been carrying at central experimental base of VNIIMK (Krasnodar) and Armavir experimental station (Armavir). Open pollinated varieties Rodnik, Master and Oreshek as well as lines VK276, VK678, VA6, VA769, VK580, VK585 and VA384 were used. No resistant plants were obtained after observation on 680000 plants of three OP varieties and seven lines grown on three experimental fields with total area of 15 ha treated with imidazolinones or sulfonylurea herbicides. DNA markers were used for molecular identification of herbicide resistance genes. DNA was extracted from the leaves of individual plants according to Saghai-Maroof methods. Particularly electrophoretic spectra of DNA fragments were estimated. It was amplificated with allele specific primers for the ImiSun gene and allele specific primers for the CLHA-Plus gene. Frequency of a dominant gene for resistance against imidazolinones was estimated less than 5 х 10 -6 (1:200000) for germplasm of OP varieties and lines of VNIIMK and less than 4х10 -6 (1 : 280000) for lines of Armavir research station. Frequency of a dominant gene for resistance against sulfonylurea was estimated less than 5 х 10 -6 (1 : 200000) for germplasm of VNIIMK lines.
Текст научной статьи Скрининг линий и сортов подсолнечника селекции ВНИИМК по устойчивости к ALS-ингибирующим гербицидам
The main task of the research was to estimate the frequency of a dominant gene for resistance against herbicides in the oil bearing germplasm of VNIIMK. Wide screening for Pulsar and Express resistance of sunflower lines and open pollinated varieties of VNIIMK breeding was done. The research has been carrying at central experimental base of VNIIMK (Krasnodar) and Armavir experimental station (Armavir). Open pollinated varieties Rodnik, Master and Oreshek as well as lines VK276, VK678, VA6, VA769, VK580, VK585 and VA384 were used. No resistant plants were obtained after observation on 680000 plants of three OP varieties and seven lines grown on three experimental fields with total area of 15 ha treated with imidazolinones or sulfonylurea herbicides. DNA markers were used for molecular identification of herbicide resistance genes. DNA was extracted from the leaves of individual plants according to Saghai-Maroof methods. Particularly electrophoretic spectra of DNA fragments were estimated. It was amplificated with allele specific primers for the ImiSun gene and allele specific primers for the CLHA-Plus gene. Frequency of a dominant gene for resistance against imidazolinones was estimated less than 5 × 10-6 (1:200000) for germplasm of OP varieties and lines of VNIIMK and less than 4×10-6 (1 : 280000) for lines of Armavir research station. Frequency of a dominant gene for resistance against sulfonylurea was estimated less than 5 × 10-6 (1 : 200000) for germplasm of VNIIMK lines.
Обнаружение в естественных популяциях дикорастущего подсолнечника растений, устойчивых к имидазолинонам и сульфонилмочевинам, вызвало сильный интерес у селекционеров, генетиков и производителей гербицидов.
Так, в штате Канзас, США на поле, обрабатываемом в течение семи лет имазе-тапиром, в популяции дикорастущего подсолнечника Helianthus annuus L. были впервые обнаружены в 1996 г. устойчивые к имидазолинонам растения [1]. В следующем 1997 г. изучали образцы дикорастущего подсолнечника, собранные с 14 полей сои, одного поля кукурузы и 11 популяций, растущих вдоль автомобильных дорог в радиусе около 20 км от исходного поля. При этом 20 популяций из 26, т.е. в 77 % случаев, содержали около 1 % устойчивых растений [2].
Дальнейшее исследование показало, что частота встречаемости полностью резистентных к имазетапиру растений в исходной популяции дикорастущего H. annuus составила 9 % при обработке 15-кратной дозой гербицида [3], а по другим данным, – 27 % при 2-кратной дозе [4]. Данная популяция использовалась в качестве источника устойчивости к имидазолиноновым гербицидам в последующих селекционно-генетических программах [5]. Признак имидазолиноно-устойчивос-ти был перенесён путём скрещивания устойчивых растений дикорастущего подсолнечника с культурными формами в гербицидоустойчивые линии НА425 и RHA426 [6].
В генофонде 29 образцов культурного подсолнечника генетической коллекции ВНИИМК, включая селекционные линии, не обнаружено генов устойчивости к имидазолиноновым гербицидам [7].
С другой стороны, создание сульфо-нилмочевино-устойчивых гибридов подсолнечника стало возможным после обнаружения также в дикорастущих популяциях резистентных особей к трибенурон-метилу. Так, 57 % из 46 природных популяций H. annuus и трёх популяций H. petiolaris Nutt., семена которых были собраны в ходе экспедиций в пяти штатах США в 2002 г., обладали устойчивыми к трибенурон-метилу особями [8]. При этом в южных регионах, где сульфонилмоче-вины использовались более длительное время, доля популяций с устойчивыми растениями была выше, например в штате Канзас достигала 94 %.
Схожие данные были получены при изучении популяционной структуры 23 образцов дикораcтущего H. annuus из хранящейся в генбанке коллекции, собранной в трёх провинциях Канады в 1994 г. Так, 52 % популяций обладали резистентными особями, а доля устойчивых к трибенурон-метилу растений в этих популяциях достигала 6 % при среднем значении 3 %. При этом шесть популяций H. petiolaris из двух провинций Канады не обладали резистентными растениями [9]. Интродукция гена устойчивости к трибенурон-метилу от дикорастущего H. annuus из Канзаса в генофонд культурного подсолнечника завершилась созданием двух доступных к использованию источников этого признака SURES-1 и SURES-2 [10].
Установлено отсутствие мутантного гена устойчивости к трибенурон-метилу у четырёх селекционных линий и 66 семей М 2 из генетической коллекции ВНИИМК [11].
В связи с отсутствием гербицидоустойчивых генотипов у культурного подсолнечника при оценке десятков растений на каждую инбредную линию или сортообразец нами была поставлена задача изучить частоту встречаемости генов гербицидоустойчивости в генофонде культурного подсолнечника ВНИИМК при существенно большем объёме выборки.
Материал и методы. Исследования проводили на центральной экспериментальной базе ВНИИМК (г. Краснодар) и Армавирской опытной станции ВНИИМК (г. Армавир). Использовали сорта-популяции Родник, Мастер и Орешек, материнские линии ВК276, ВК678, ВА6 и ВА769, отцовские линии ВК580, ВК585 и ВА384. Посев сеялкой проводили на трёх 5-гектарных участках: первом (ЦЭБ) в 2012 г., втором (ЦЭБ) и третьем (АОС) – в 2013 г. В фазе трех пар настоящих листьев растения обрабатывали гербицидами Пульсар (1 л/га, д.в. имазамокс) на участках № 1 и № 3, а также гербицидом
Экспресс (35 г/га, д.в. трибенурон-метил) на участке № 2. Оценку поражения растений гербицидом проводили через семь дней после обработки. Явно ложноустойчивые типичные растения в пределах ряда, т.е. группы растений, на которые не попал гербицид, а также и единичные поздно взошедшие растения, избежавшие контакта с гербицидом, не учитывались как устойчивые.
ДНК подсолнечника выделяли из листьев индивидуальных вегетирующих растений. Выделение проводили по модифицированному методу, описанному Saghai-Maroof с соавторами [12].
Для проведения полимеразной цепной реакции использовали 25 мкл реакционной смеси следующего состава: 67 мМ трис-HCl, рН 8,8; 16,6 мM сульфата аммония; 1,5–3 мM MgCl 2 ; 0,01 % Tween 20; по 0,2 мM дезоксирибонуклеозидфосфатов; по 20 мкМ праймеров; 5Х Q solution ; 10 нг матричной ДНК и 1 ед. рекомбинантной термостабильной ДНК полимеразы (Москва, Сибэнзим). Для амплификации использовали термоциклер S1000тм (BioRad, США). Условия амплификации: начальная денатурация – 2 мин 96 оС, затем 30 циклов при соблюдении температурно-временного режима: денатурация при 94 °С – 30 сек, отжиг при 60 °С в течение 40 сек, элонгация – 1 мин при 70 °С, финальная элонгация – 2 мин. Для ПЦР анализа использовали SNP-праймеры, которые были разработаны и любезно предоставлены нам фирмой BASF (США).
Электрофорез продуктов амплификации проводили в агарозном геле (2 % агароза, 1х ТАЕ-буфер) с использованием камеры SE-2 для горизонтального электрофореза (Хеликон, РФ). Последующее окрашивание осуществляли бромистым этидием. Документирование результатов электрофореза обеспечивалось при помощи системы цифровой документации видеоизображения BIO-PRINT (Vilber Lourmat, Франция).
Результаты и обсуждение. В опыте 2012 г. на поле № 1, обработанном Пульсаром, были обнаружены семь растений без признаков угнетенности, тогда как все остальные из около 200000 осмотренных растений сортов Родник, Мастер и Орешек, а также линий ВК678 и ВК580 обладали явными симптомами повреждения гербицидом. Особи без признаков поражения гербицидом были выкопаны, пересажены и самоопылены в питомнике на селекционном поле.
Семь полученных инбредных семей были оценены по устойчивости к Пульсару в условиях камеры фитотрона в осеннезимний период 2012–2013 гг. (табл. 1). Шесть семей оказались повреждёнными гербицидом (рис.1), следовательно, исходные растения были ложно устойчивыми. Однако одна семья от родоначального растения из ряда линии ВК580 в полевом посеве, показала моногенное расщепление на устойчивые и погибшие (рис. 2) в отношении 14 : 4, χ2 3:1 = 0,07, χ2 st = 3,84, р > 0,05 при доминировании устойчивости. Расщепление в I 1 указало на гетерозиготность родоначального растения. С выращенных особей с максимальной степенью устойчивости, обладающих при этом ветвистым фенотипом, были получены семена I 2 в условиях камеры фитотрона.
Таблица 1
Оценка семи отобранных в поле семей I 1 по устойчивости к Евро-Лайтнингу (доза 0,25х ) в камере фитотрона
ЦЭБ ВНИИМК, Краснодар, 2012 г.
|
Генотип |
Число растений I 1 , шт. |
|
|
устойчивые |
погибшие |
|
|
Мастер |
0 |
2 |
|
Родник |
0 |
12 |
|
Родник |
0 |
6 |
|
Орешек |
0 |
16 |
|
ВК 678 |
0 |
17 |
|
ВК 580 |
0 |
11 |
|
ВК «580» |
14 |
4 |

Рисунок 1 – Погибшие растения семьи I 1 ВК580 через шесть дней после обработки Евро-Лайтнингом (доза 0,25х)

Рисунок 2 – Расщепление по гербицидо-устойчивости растений в семье I 1 ВК580 через шесть дней после обработки
Евро-Лайтнингом (доза 0,25х)
В полевых условиях 2013 г. все растения одной из отобранных семей I 2 ВК580 оказались одновременно ветвистыми и устойчивыми к имидазолиноновому гербициду Евро-Лайтнинг, т.е. были гомозиготны по генам этих признаков. Однако по комплексу морфологических признаков растения этой семьи однозначно отличались от ожидаемого морфотипа линии ВК580. Поскольку на скрининговом поле № 1 в 2012 г. была посеяна только одна ветвистая линия ВК580, то очевидно, что родоначальное гербицидоустойчивое гетерозиготное растение, обнаруженное в ряду растений ВК580, было или падалицей, или ауткроссом в предыдущих поколениях (т.е. условно «ВК580»).
Более того, молекулярно-генетическая идентификация с помощью SNP-маркеров, проведённая совместно с лабораторией иммунитета и электрофореза ВНИИМК, показала аллельность обнаруженной мутации гену Imr , уже используемому в селекции и сельскохозяйственном производстве.
Был проведен анализ образцов подсолнечника гебицидоустойчивой семьи «ВК580» по маркерам, выявляющим точечные мутации в разных позициях гена устойчивости к имидазолинонам AHASL1. Дорожки 1–3 на рисунке 3 являются контрольными и показывают образцы подсолнечника, несущие мутацию A205 (At)V. Фракция длиной 970 пн представляет собой контрольный бенд, который амплифицируется несмотря на наличие или отсутствие мутации. Фрагмент ДНК 666 пн является диагностическим для му- тантного генотипа. И, наконец, фрагмент 304 пн амплифицируется у образцов подсолнечника дикого типа, т.е. не несущего мутацию A205 (At)V. Дорожки 4–8 на рис. 3 – ДНК образцов подсолнечника, которые были толерантны к имидазолиноновым гербицидам в поле, но природа устойчивости была неизвестна. Поскольку у этих образцов амплифицировался диагностический бенд 666 пн, но не ам-плифицировался фрагмент 304 пн, данные образцы подсолнечника являются гомозиготными мутантами по гену ImiSun (или Imr).
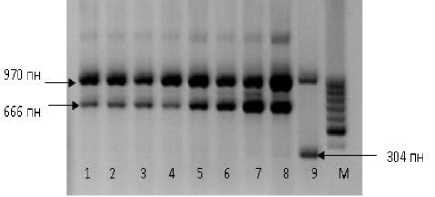
Рисунок 3 – Электрофоретические спектры фрагментов ДНК подсолнечника, ампли-фицированные с аллель-специфичными праймерами по гену ImiSun .
М – маркер молекулярного веса 100 kb. Дорожки: 1–3 – ДНК образцов подсолнечника, несущих мутацию A205 (At)V; 4–8 – ДНК образцов подсолнечника, толерантных к имидазолиноновым гербицидам; 9 – ДНК чувствительного подсолнечника дикого типа, неустойчивого к имидазолинону
Дорожки 1–5 на рисунке 4 представляют образцы подсолнечника, несущие мутацию A122 (At)T гена AHASL1 . Фрагмент ДНК 576 пн является диагностическим для этого мутантного генотипа. Фрагмент 258 пн амплифицируется у образцов подсолнечника дикого типа. Дорожки 6–10 на рисунке 2 – ДНК образцов подсолнечника, которые были толерант-ны к имидазолиноновым гербицидам, но природа устойчивости была неизвестна. Поскольку у этих образцов не амплифици-ровался диагностический бенд 576 пн, но амплифицировался фрагмент 258 пн, установлено, что данные образцы подсолнечника не несут мутации по сайту A122 (At)T гена AHASL1 , т. е. гену CLHA-Plus .
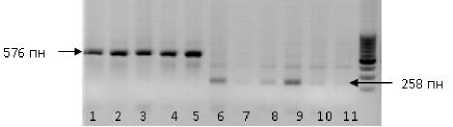
Рисунок 4 – Электрофоретические спектры фрагментов ДНК подсолнечника, ампли-фицированные с аллель-специфичными праймерами по гену CLHA-Plus .
М – маркер молекулярного веса 100 kb. Дорожки: 1–5 ДНК образцов подсолнечника, несущих мутацию A122 (At)T гена AHASL1 ; 6–10 – ДНК образцов подсолнечника, которые были толерантны к имидазолиноновым гербицидам; 11 – контрольный образец ДНК подсолнечника дикого типа, нечувствительный к имидазолинону
В 2013 г. на поле № 2 площадью 5 га около 200000 растений линий ВК276, ВК678, ВК580 и ВК585 были обработаны гербицидом Экспресс (1х доза) для поиска устойчивых к сульфонилмочевинам генотипов. После оценки фитотоксичности пять растений без признаков повреждения гербицидом были выкопаны и пересажены в селекционный питомник для получения следующего поколения. Четыре растения по комплексу морфологических признаков явно не соответствовали по фенотипу ни одной из посеянных линий. Потомство одного растения, фенотипически похожего на ВК276, при выращивании в камере фитотрона в осеннезимний период 2013–2014 гг. показало отсутствие гербицидоустойчивости. Очевидно, что родоначальное растение линии ВК276 без видимых признаков повреждения гербицидом на поле № 2 было ложно устойчивым.
В результате совместного опыта с Армавирской опытной станцией ВНИИМК на 5-гектарном поле подсолнечника № 3 после обработки Пульсаром (1 л/га) были отобраны, выращены и самоопылены два растения, по одному из рядов линий ВА6 и ВА384 в посеве, без признаков повреждения гербицидом. В камере фитотрона в осенне-зимний период 2013–2014 гг. семья I 1 «ВА6» показала моногенное расщепление по гербицидоустойчивости, тогда как все растения семьи I 1 ВА384
полностью погибли, показав ложную устойчивость родоначального растения. Выжившие растения семьи I 1 «ВА6» по комплексу морфологических признаков, включая окраску и форму как листьев, так и околоплодника семян, явно не соответствовали фенотипу линии ВА6, что указывает на родоначальное растение как падалицу или ауткросс в предыдущих поколениях.
Таким образом, на трёх экспериментальных полях общей площадью 15 га при осмотре около 680000 растений трёх сортов и семи линий не было обнаружено ни одного типичного растения посеянных генотипов подсолнечника с признаком устойчивости к ALS-ингибирующим гербицидам: имидазолинонам или сульфо-нилмочевинам (табл. 2).
Таблица 2
Оценка частоты встречаемости доминантных генов гербицидоустойчивости в селекционном материале подсолнечника ВНИИМК
ВНИИМК, 2012–2013 гг.
|
Генотип |
Количество обработанных растений, шт. |
Количество устойчивых растений, шт. |
Частота встречаемости генов гербицидо-устойчивости |
|
Поле №1, 200000 растений, Пульсар, 2012 г. |
|||
|
Родник |
20000 |
0 |
< 1/20000 |
|
Мастер |
20000 |
0 |
< 1/20000 |
|
Орешек |
20000 |
0 |
< 1/20000 |
|
ВК276 |
30000 |
0 |
< 1/30000 |
|
ВК678 |
30000 |
0 |
< 1/30000 |
|
ВК580 |
40000 |
0 |
< 1/40000 |
|
ВК585 |
40000 |
0 |
< 1/40000 |
|
Поле №2, 200000 растений, Экспресс, 2013 г. |
|||
|
ВК276 |
40000 |
0 |
< 1/40000 |
|
ВК678 |
40000 |
0 |
< 1/40000 |
|
ВК580 |
60000 |
0 |
< 1/60000 |
|
ВК585 |
60000 |
0 |
< 1/60000 |
|
Поле №3, 280000 |
растений, Пульсар, 2013 г. |
||
|
ВА6 |
100000 |
0 |
< 1/100000 |
|
ВА760 |
70000 |
0 |
< 1/70000 |
|
ВА384 |
110000 |
0 |
< 1/110000 |
Заключение. Частота встречаемости доминантного гена имидазолиноно-устой-чивости оценивается менее чем 5 × 10-6 (1 : 200000) для генофонда изученных сортов и линий ЦЭБ ВНИИМК и менее 4 × 10-6 для линий Армавирской опытной станции (1 : 280000). Частота встречаемости доминантного гена устойчивости к трибенурон-метилу у изученных селекционных линий не превышает значения 5 × 10-6 (1:200000).
