Создание и изучение рекомбинантных инбредных линий подсолнечника с различным содержанием олеиновой кислоты в масле семян

Автор: Демурин Я.Н., Чебанова Ю.В., Борисенко О.М., Гучетль С.З., Савиченко Д.Л., Ефименко С.Г., Рубанова О.А., Широких А.А., Шамрай В.Д.
Рубрика: Селекция и семеноводство сельскохозяйственных растений
Статья в выпуске: 2 (182), 2020 года.
Бесплатный доступ
Создание и изучение рекомбинантных инбредных линий является эффективным методом в селекционно-генетической работе с полигенными признаками. В нашем исследовании подобный подход использован для признака содержания олеиновой кислоты в масле семян подсолнечника. Опыты проводили в полевых и лабораторных условиях во ВНИИМК, г. Краснодар в период 2012-2020 гг. Для получения рекомбинантных инбредных линий с различным количеством олеиновой кислоты в масле семян проводили скрещивание, самоопыление и дизруптивный отбор для трех реципрокных комбинаций ЛГ27 × ЛГ28, ЛГ27 × ВК678 и ЛГ27 × ЛГ26. Линия ЛГ26 обладает высокоолеиновым фенотипом, ЛГ27 - среднеолеиновым, ВК678 - повышенноолеиновым и ЛГ28 - низкоолеиновым. Анализ жирно-кислотного состава масла семян подсолнечника проводили с использованием метода газожидкостной хроматографии метиловых эфиров на приборах Хром-5 и Хроматэк-Кристалл 5000. Молекулярно-генетическое маркирование высокоолеиновых образцов подсолнечника проводили с использованием двух пар праймеров: F13/R5 и F4/R1...
Олеиновая кислота, линия, скрещивание, молекулярный маркер, мутация
Короткий адрес: https://sciup.org/142223426
IDR: 142223426 | УДК: 633.854.78:575 | DOI: 10.25230/2412-608X-2020-2-182-3-12
Development and study of recombinant inbred lines of sunflower with different content of oleic acid in seed oil
The creation and study of recombinant inbred lines is an effective method in breeding and genetic work with polygenic traits. In our study, a similar approach was used to indicate the content of oleic acid in sunflower seed oil. The experiments were carried out in the field and laboratory conditions in VNIIMK, Krasnodar, in the period 2012-2020. To obtain recombinant inbred lines with different contents of oleic acid in seed oil, hybridization, self-pollination and disruptive selection were used for three reciprocal combinations LG27 × LG28, LG27 × VK678 and LG27 × LG26. The LG26 line has a high oleic phenotype, LG27 has a mid-oleic, VK678 has an increased oleic and LH28 has a low oleic. The analysis of the fatty acid composition of sunflower seed oil was carried out using the method of gas-liquid chromatography of methyl esters on Chrom-5 and Chromatek-Crystal 5000 instruments. Molecular genetic marking of high oleic sunflower samples was performed using two pairs of primers: F13/R5 and F4/R1...
Текст научной статьи Создание и изучение рекомбинантных инбредных линий подсолнечника с различным содержанием олеиновой кислоты в масле семян
Создание и изучение рекомбинантных инбред-ных линий является эффективным методом в селекционно-генетической работе с полигенными признаками. В нашем исследовании подобный подход использован для признака содержания олеиновой кислоты в масле семян подсолнечника. Опыты проводили в полевых и лабораторных условиях во ВНИИМК, г. Краснодар в период 2012– 2020 гг. Для получения рекомбинантных инбред-ных линий с различным количеством олеиновой кислоты в масле семян проводили скрещивание, самоопыление и дизруптивный отбор для трех реципрокных комбинаций ЛГ27 × ЛГ28, ЛГ27 × ВК678 и ЛГ27 × ЛГ26. Линия ЛГ26 обладает вы-сокоолеиновым фенотипом, ЛГ27 – среднеолеи-новым, ВК678 – повышенноолеиновым и ЛГ28 – низкоолеиновым. Анализ жирно-кислотного состава масла семян подсолнечника проводили с использованием метода газожидкостной хроматографии метиловых эфиров на приборах Хром-5 и Хроматэк-Кристалл 5000. Молекулярно генетическое маркирование высокоолеиновых образцов подсолнечника проводили с использованием двух пар праймеров: F13/R5 и F4/R1. Исходные родительские линии ЛГ28, ВК678, ЛГ27 и ЛГ26 подтвердили принадлежность к своим фенотипическим классам. При этом с увеличением содержания олеиновой кислоты с 29 до 88 % наблюдалось существенное снижение значений показателя изменчивости признака в пределах классов CV c 14 до 1 %. Созданные рекомбинантные инбредные линии подсолнечника F5 при скрещиваниях среднеолеиновой ЛГ27 охватывают широкий интервал значений содержания олеиновой кислоты (от 30 до 92 %) в масле семян и являются ценным материалом для дальнейших исследований. Доказана полная эффективность использования молекулярного маркера F4/R1 в идентификации мутации высокоолеиновости у линий с высоким содержанием олеиновой кислоты в фенотипическом интервале 87–92 %. При этом полученные среднеолеиновые рекомбинантные инбредные линии и ЛГ27 не обладают мутацией Ol.
UDC 633.854.78:575
Development and study of recombinant inbred lines of sunflower with different content of oleic acid in seed oil.
Ya.N. Demurin, doctor of biology
Yu.V. Chebanova, PhD in biology
O.M. Borisenko, PhD in biology
S.Z. Guchetl, PhD in biology
D.L. Savichenko, researcher
S.G. Efimenko, PhD in biology
O.A. Rubanova, post-graduate student
A.A. Shirokikh, Master’ degree student
-
V.D. Shamray, schoolgirl
-
V.S. Pustovoit All-Russian Research Institute of Oil Crops (VNIIMK)
-
17, Filatova str., Krasnodar, 350038, Russia
The creation and study of recombinant inbred lines is an effective method in breeding and genetic work with polygenic traits. In our study, a similar approach was used to indicate the content of oleic acid in sunflower seed oil. The experiments were carried out in the field and laboratory conditions in VNIIMK, Krasnodar, in the period 2012–2020. To obtain recombinant inbred lines with different contents of oleic acid in seed oil, hybridization, selfpollination and disruptive selection were used for three reciprocal combinations LG27 × LG28, LG27 × VK678 and LG27 × LG26. The LG26 line has a high oleic phenotype, LG27 has a mid-oleic, VK678 has an increased oleic and LH28 has a low oleic. The analysis of the fatty acid composition of sunflower seed oil was carried out using the method of gasliquid chromatography of methyl esters on Chrom-5 and Chromatek-Crystal 5000 instruments. Molecular genetic marking of high oleic sunflower samples was performed using two pairs of primers: F13/R5 and F4/R1. The initial parental lines LG28, VK678, LG27 and LG26 confirmed their belonging to their phenotypic classes. At the same time, with an increase in the oleic acid content from 29 to 88%, a significant decrease in the values of variability of the trait within classes CV from 14 to 1% was observed. The created recombinant inbred lines of sunflower F 5 for crossing mid-oleic LG27 cover a wide range of oleic acid content from 30 to 92% in seed oil and are valuable material for further research. The full efficiency of using the molecular marker F4/R1 in the identification of high oleic mutations in lines with high oleic acid content in the phenotypic range of 87–92% has been proved. In this case, the mid-oleic LG27 and the mid-oleic developed recombinant inbred lines do not possess the Ol mutation.
Введение. Начиная с 1976 г., когда во ВНИИМК впервые в мире был создан вы-сокоолеиновый сорт подсолнечника Первенец [1], инициируется новый этап в селекции подсолнечника на качество масла. В первоначальных работах по изучению генетического контроля высоко-олеиновости установлено, что этот признак контролируется одним доминантным геном Ol [2] либо одним частично доминантным геном [3]. Последующие исследования указали на присутствии гена-модификатора Ml, гомозиготное рецессивное состояние которого позволяет полностью проявляться мутации высокоолеиновости, тогда как генотип OlolMlml дает промежуточный фенотипический класс с содержанием олеиновой кислоты около 48–72 % [4; 5]. Fernandez-Martinez et al. обнаружили, что признак высокоолеиновости контролировался тремя доминантными комплементарными генами Ol1, Ol2 и Ol3 [6], позже данную гипотезу подтвердили Perez-Vich et al. [7]. В ходе дальнейших исследований Velasco et al. предложили гипотезу о пяти комплементарных генах, регулирующих содержание олеиновой кислоты (Ol1, Ol2, Ol3, Ol4 и Ol5) [8].
Во ВНИИМК в результате проведенного гибридологического анализа установлено, что признак высокоолеиновости контролируется одним доминантным геном Ol с неполной пенетрантностью в гетерозиготе [9]. Кроме того, было обнаружено наличие супрессора, обуславливающего неполное проявление гена Ol в гетерозиготном состоянии [10]. У низкоолеиновой линии RIL100, созданной во Франции, доказано наличие мутации Ol в гипостатическом состоянии за счет влияния генов-супрессоров. Обнаружено также, что линия ЛГ26 характеризуется устойчивостью мутации Ol к действию супрессора [11].
В настоящее время большое внимание уделяется изучению молекулярных причин изменения содержания жирных кислот в масле семян подсолнечника. Lacombe et al. обнаружили, что высокое содержание олеиновой кислоты в сорте Первенец контролировалось двумя независимыми локусами, первый представлен oleHOS аллелем, а другой – FAD2-1 аллелем [12]. Hongtrakul et al. объединяют ген Ol с геном OLD-7 (FAD 2-1), проявление которого слабо выражено у высокоолеи-новых генотипов, но сильно выражено у обычных генотипов [13]. Pérez-Vich et al. установили, что QTL (qualitative trait loci – локусы количественного признака, ЛКП) для олеиновой кислоты расположены в локусе OLD-7 (14-я группа сцепления) и предположили, что он является модификатором Ol [7]. Premnath et al. также картировали ген Ol в 14-й группе сцепления (LG) и связали с маркером HO_Fspb.
Кроме того, в группах сцепления LG8 и LG9 идентифицировано еще два QTL для содержания олеиновой кислоты [14].
Schuppert et al. показали, что мутация Ol коррелирует со значительно сниженной экспрессией FAD2-1. Было обнаружено, что мутант несет тандемные повторы FAD2-1, разделенные межгенной областью. Эти ученые разработали кодоминантные маркеры SSR и INDEL для FAD2-1 с целью диагностики наличия мутации Ol [15].
В Индии провели изучение двух линий и их гибрида с использованием молекулярных маркеров SSR и INDEL, которые связаны с высоким содержанием олеиновой кислоты. Результаты позволили проверить один SSR (N1-3F/N1-3R) и два INDEL (F4/R1 и F4/R2) маркера в генотипах подсолнечника для идентификации признака высокого содержания олеиновой кислоты [16]. В Сербии протестировали несколько молекулярных маркеров для ускорения селекции при обнаружении высокоолеиновых генотипов. Было доказано, что маркер F4/R1 наиболее эффективен при идентификации генотипов с мутацией Ol в поколении F 2 [17].
Изучению признака среднеолеиново-сти уделялось относительно меньше внимания. В 1999 г. в USDA (г. Фарго, США) были выведены среднеолеиновые линии подсолнечника HA421, HA422 и HA424, содержащие около 63–67 % олеиновой кислоты в масле семян [18]. Однако сред-неолеиновый фенотип этих линий связан с гетерозиготностью и расщеплением по мутации высокоолеиновости [19], тогда как в генетической коллекции подсолнечника ВНИИМК содержится линия ЛГ27 с гомозиготным константным среднеолеи-новым фенотипом [20]. Первоначально во ВНИИМК была высказана гипотеза о том, что признак повышенного содержания олеиновой кислоты линии ЛГ27 контролируется рецессивным аллелем, обозначенным ol′, отличающимся как от мутации Ol, так и от аллеля дикого типа ol [20]. Позже было доказано, что признак среднеолеиновости ЛГ27 находится под контролем аддитивной олигогенной системы с материнским эффектом в F1 и отсутствием материнского наследования в F2 и F3 [21; 22]. Однако молекулярногенетического тестирования на наличие мутации Ol у линии ЛГ27 не проводилось.
В 1999 г. в США в соответствии с запросом пищевой промышленности было выпущено на рынок среднеолеиновое подсолнечное масло NuSun [23]. Высоко-олеиновые гибриды подсолнечника выращиваются в нашей стране [24; 25], а создание и коммерциализация средне-олеиновых гибридов находятся на начальном этапе развития. Однако в других странах мира, например, США и Канаде, среднеолеиновые гибриды подсолнечника уже около 20 лет составляют основную долю посевных площадей этой культуры. По международной классификации типов подсолнечного масла CODEX Stan 210 традиционное масло содержит 14–39 % олеиновой кислоты, среднеолеиновое – 43–72 %, а высокоолеиновое – 75–91 % [26]. Главное преимущество среднеолеи-нового масла в том, что оно обладает оптимальным сочетанием высокой окислительной стабильности и достаточным содержанием эссенциальной линолевой кислоты около 20 %.
Создание и изучение рекомбинантных инбредных линий (РИЛ, RIL) является эффективным методом в селекционногенетической работе с разными признаками, особенно с полигенными. Цель данного исследования заключалась в использовании подобного подхода к признаку содержания олеиновой кислоты в масле семян подсолнечника.
Материалы и методы. Опыты проводили в полевых и лабораторных условиях во ВНИИМК, г. Краснодар, в период 2012–2020 гг. Агроклиматические условия, сложившиеся в период проведения полевых исследований, можно считать удовлетворительными для изучаемой культуры. Закладку опытов, включая скрещивания и принудительное самоопыление растений, выполняли согласно общепринятой методике.
Использовали константные линии генетической коллекции подсолнечника ВНИИМК: высокоолеиновую – ЛГ26; среднеолеиновую – ЛГ27; повышенно-олеиновую – ВК678; низкоолеиновую – ЛГ28. Кроме того, линия ЛГ26 обладает рядом морфологических особенностей, в частности оранжевыми узкояйцевидными язычковыми цветками и оранжевыми с антоцианом в отгибе венчика трубчатыми цветками.
Анализ жирно-кислотного состава масла семян подсолнечника проводили в лаборатории биохимии с использованием метода газожидкостной хроматографии метиловых эфиров на приборах Хром-5 и Хроматэк-Кристалл 5000. Статистическую обработку данных проводили в программе Microsoft Excel 2010.
Геномную ДНК выделяли из семядольных листьев 5–7-дневных этиолированных проростков подсолнечника с помощью модифицированного метода Saghai-Maroof et аl. [27] с использованием СТАВ буфера. Для проведения ПЦР использовали 25 мкл реакционной смеси следующего состава: 67 мМ трис-HCl, рН 8,8; 16,6 мM сульфата аммония; 1,5–3 мM MgCl2; 0,01 % Tween 20; по 0,2 мM дезоксирибонуклеозидфосфатов; по 10 пМ праймеров; 10 нг матричной ДНК и 1 ед. рекомбинантной термостабильной ДНК полимеразы (Сибэнзим, Москва). Амплификацию выполняли в термоциклере S1000тм (BioRad, США). Условия амплификации: начальная денатурация при 96 °С в течение 2 мин.; затем 30 циклов при соблюдении температурно-временного режима: отжиг при 60 °С в течение 40 сек, элонгация – 1 мин при 70 °С, денатурация при 94 °С – 30 сек, финальная элонгация – 2 мин. Использовали четыре INDEL праймера F4, F13, R1, R5, разработанных Schuppert et al. [15]. Праймеры были изготовлены ООО «Синтол» (Москва, Россия). Электрофорез продуктов ампли- фикации проводили в агарозном геле (2 % агароза, 1 х SB-буфер) с использованием камеры для горизонтального электрофореза (SE.2, ДНК-технология, Россия) в течение 1–1,5 часов при силе тока 58 mA и напряжении 90–100 V. Последующее окрашивание осуществляли бромистым этидием. Визуализация результатов электрофореза в ультрафиолетовом свете и их документирование обеспечивали при помощи системы цифровой документации видеоизображения BIO-PRINT (Vilber Lourmat, Франция). Размер фрагментов ДНК определяли с использованием программного обеспечения Bio-Capture (Vilber Lourmat, Франция) относительно маркера длины фрагментов ДНК GeneRuler 100 bp DNA Ladder Thermo Scientific (Сибэнзим, Россия).
Результаты и обсуждение . Для получения рекомбинантных инбредных линий с различным количеством олеиновой кислоты в масле семян проводили скрещивание, самоопыление и дизруптивный отбор для трех реципрокных комбинаций ЛГ27 × ЛГ28, ЛГ27 × ВК678 и ЛГ27 × ЛГ26 (табл. 1).
В 2012 г. были получены семена F 1 . После самоопыления растений в 2013 г. сформировались семена F 2 (I 1 F 1 ), которые были индивидуально проанализированы с сохранением жизнеспособности зародыша. От выращенных в 2014 г. растений из контрастных фенотипических классов получены инбредные семена I 2 с минимальным и максимальным проявлением признака. На каждой делянке самоопыля-ли по 5 растений. Семена поколения I 3 получили в 2015 г. без проведения анализов. В 2016 г. отобрали 145 самоопылен-ных корзинок поколения I 4 (линии F 5 ) от всех реципрокных скрещиваний (табл. 1). Кроме того, в 2019 г. для части линий проведено очередное самоопыление.
В поколении I 4 от скрещивания ЛГ28 и ЛГ27 выделили 46 линий с содержанием олеиновой кислоты в пределах 29,8–56,9 %. Из них 14 вошли в обычный (30–40 % С 18:1 ), 31 – в повышенноолеиновый (41– 54 %), одна линия (56,9 %) – в средне-олеиновый фенотипический класс (табл. 1, табл. 2).
Таблица 1
Рекомбинантные линии от реципрокных скрещиваний линий ЛГ27 и ВК678 в количестве 57 штук содержали от 36,5 до 64,8 % олеиновой кислоты. Из них пять линий относились к обычному классу (30–40 % С 18:1 ), 35 – к повышенноолеино-вому (41–54 %), 17 – к средне-олеиновому (55–65 %) (табл. 1, табл. 2).
Гибридизация ЛГ27 и ЛГ26 дала 42 рекомбинантные линии I 4 с большим интервалом варьирования (до 50 %) по содержанию олеиновой кислоты. Эти линии вошли в четыре фенотипических класса: две – в обычный класс (30–40 % С 18:1 ), 13 – в повышенноолеиновый (41– 54 %), 10 – в среднеолеиновый (55–77 %) и 11 – в высокоолеиновый (86–92 %). Из них семь линий являются супер-высокоолеиновыми (90–92 % С 18:1 ), т.е. за счет положительной трансгрессии получены генотипы с проявлением признака выше, чем у родительской высокоолеино-вой линии ЛГ26 (табл. 1, табл. 2).
Родительские линии ЛГ28, ВК678, ЛГ27 и ЛГ26 подтвердили свой низко-, повышено-, средне- и высокоолеиновый фенотип соответственно (табл. 2). При этом с увеличением содержания олеиновой кислоты с 29,3 до 87,9 % наблюдается очевидное снижение значений показателя изменчивости признака CV c 14 до 1 %.
Молекулярное маркирование высоко-олеиновых образцов подсолнечника проводили с использованием двух пар праймеров: F13/R5 и F4/R1. Пара праймеров F13/R5 в результате ПЦР гибридизу-ется с ДНК как высокоолеинового, так и линолевого фенотипа, то есть происходит амплифицикация участка ДНК, несмотря на наличие или отсутствие мутации Ol [15]. Данный локус использовался как положительный контроль для исключения ложноотрицательного результата на наличие маркируемой мутации. Испытываемые образцы в результате ПЦР ДНК 8
выявили фракцию размером около 340 п. н., характеризующую как мутантный, так и нормальный генотипы (рис. 1, табл. 3).
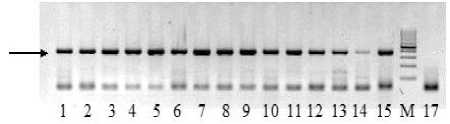
Рисунок 1 – Электрофоретические спектры ДНК образцов подсолнечника при амплификации с праймерами F13/R5
Примечание : дорожки 1–3 – № 13; 4–6 – № 3; 7–9 – № 8; 10–12 – № 7; 13–15 – № 21;
М – маркер молекулярного веса 100 п. н.;
17 – отрицательный контроль. Стрелкой обозначен фрагмент ДНК 340 п. н., присутствующий у всех генотипов (положительный контроль).
№ образца из таблицы 3
Доминантный INDEL маркер F4/R1 разработан для диагностики наличия мутации Ol у подсолнечника [15]. В испытываемой коллекции RIL пара праймеров F4/R1 амплифицировала фракцию ДНК примерно 590 п. н. у высокоолеиновых образцов. Эта полоса отсутствовала в образцах подсолнечника с низкоолеиновым фенотипом (рис. 2, табл. 3).
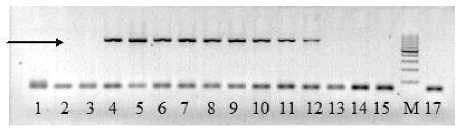
Рисунок 2 – Электрофоретические спектры ДНК образцов подсолнечника при амплификации с праймерами F4/R1. Примечание : дорожки 1–3 – № 13; 4–6 – № 3; 7–9 – № 8; 10–12 – № 7; 13–15 – № 21; М – маркер молекулярного веса 100 п. н.;
17 – отрицательный контроль. Стрелкой обозначен фрагмент ДНК 590 п. н., маркирующий мутацию высокоолеиновости масла. № образца из таблицы 3
Таблица 3
Наличие мутации Ol в рекомбинантных инбредных линиях подсолнечника
ВНИИМК, г. Краснодар, 2020 г.
|
№ |
Генотип |
Содержание C 18:1 , % |
Маркер F4/R1 мутации Ol |
|
1 |
F 6 ЛГ26 × ЛГ27 |
92,2 |
+ |
|
2 |
F 6 ЛГ26 × ЛГ27 |
92,2 |
+ |
|
3 |
F 6 ЛГ26 × ЛГ27 |
92,0 |
+ |
|
4 |
F 6 ЛГ26 × ЛГ27 |
91,3 |
+ |
|
5 |
F 6 ЛГ26 × ЛГ27 |
91,3 |
+ |
|
6 |
F 6 ЛГ26 × ЛГ27 |
91,1 |
+ |
|
7 |
F 6 ЛГ26 × ЛГ27 |
90,6 |
+ |
|
8 |
F 6 ЛГ26 × ЛГ27 |
89,5 |
+ |
|
9 |
F 6 ЛГ26 × ЛГ27 |
89,4 |
+ |
|
10 |
F 6 ЛГ27 × ЛГ26 |
88,5 |
+ |
|
11 |
F 6 ЛГ27 × ЛГ26 |
87,6 |
+ |
|
12 |
F 6 ЛГ26 × ЛГ27 |
86,5 |
+ |
|
13 |
F 6 ЛГ26 × ЛГ27 |
65,5 |
- |
|
14 |
F 6 ЛГ26 × ЛГ27 |
62,8 |
- |
|
15 |
F 6 ЛГ26 × ЛГ27 |
58,8 |
- |
|
16 |
F 6 ЛГ26 × ЛГ27 |
58,5 |
- |
|
17 |
F 6 ЛГ26 × ЛГ27 |
58,2 |
- |
|
18 |
F 6 ЛГ26 × ЛГ27 |
55,6 |
- |
|
19 |
F 6 ЛГ26 × ЛГ27 |
53,4 |
- |
|
20 |
F 6 ЛГ27 × ЛГ26 |
48,3 |
- |
|
21 |
F 6 ЛГ27 × ЛГ26 |
47,5 |
- |
|
22 |
F 6 ЛГ27 × ЛГ26 |
47,0 |
- |
|
23 |
F 6 ЛГ27 × ЛГ26 |
42,1 |
- |
|
24 |
F 6 ЛГ27 × ЛГ26 |
40,9 |
- |
|
25 |
F 6 ЛГ27 × ЛГ26 |
39,8 |
- |
|
26 |
F 6 ЛГ27 × ЛГ26 |
39,8 |
- |
|
27 |
F 6 ЛГ27 × ЛГ26 |
39,0 |
- |
|
ЛГ26, high oleic ЛГ27, mid oleic |
88,2 65,9 |
+ - |
С помощью вышеперечисленных пар праймеров в 2020 г. проанализировано, в сумме, 27 рекомбинантных инбредных линий подсолнечника F 6 (I 5 F 1 ) урожая 2019 г. Молекулярный маркер F13/R5, используемый в качестве положительного контроля, показал наличие контрольной фракции ДНК у всех анализируемых линий (табл. 3). Одновременно с этим при проведении каждой ПЦР использовался отрицательный контроль для исключения ложноположительного результата. Для этого к реакционной смеси добавляли H 2 O, не содержащую ДНК. Таким образом, минимизировались как ложноположительные, так и ложноотрицательные результаты анализа.
Из 27 рекомбинантных инбредных линий мутация Ol , маркируемая локусом F4/R1, была выявлена у 12 с содержанием олеиновой кислоты в масле семян от 86,5
до 92,2 %. В оставшихся 15 линиях с содержанием олеиновой кислоты 39,0– 65,5 % фракция ДНК, маркирующая мутацию Ol, не обнаружена. При этом из вышеуказанных 15 линий шесть (№ 13– 18) относятся с среднеолеиновым. Родительская высокоолеиновая линия ЛГ26, как и следовало ожидать, показала наличие молекулярного маркера мутации, а среднеолеиновая ЛГ27 характеризовалась его отсутствием (табл. 3). Таким образом, с использованием молекулярных маркеров было доказано присутствие в изученных генотипах мутации Ol, которая обусловливает высокое содержание олеиновой кислоты в масле семян подсолнечника.
Выводы. Созданные рекомбинантные инбредные линии подсолнечника F 5 при скрещивании среднеолеиновой ЛГ27 с высокоолеиновой ЛГ26, повышенноолеи-новой ВК678 и низкоолеиновой ЛГ28 охватывают широкий интервал значений содержания олеиновой кислоты от 30 до 92 % в масле семян и являются ценным материалом для дальнейших исследований, включая молекулярно-генетические подходы. Доказана полная эффективность использования молекулярного маркера F4/R1 в идентификации мутации высоко-олеиновости у линий с высоким содержанием олеиновой кислоты в фенотипическом интервале 87–92 %. При этом полученные среднеолеиновые рекомбинантные инбредные линии и ЛГ27 не обладают мутацией Ol .
Список литературы Создание и изучение рекомбинантных инбредных линий подсолнечника с различным содержанием олеиновой кислоты в масле семян
- Солдатов К.И., Воскобойник Л.К., Харченко Л.Н. Высокоолеиновый сорт подсолнечника Первенец // Бюл. НТИ по масличным культурам. - 1976. - Вып. 3. - С. 3-7.
- Urie A.L. Inheritance of very high oleic acid content in sunflower // Proc. of Sunflower Research Forum. - National Sunflower Association, Bismarck, ND, 1984. - P. 9-10.
- Fick G.N. Inheritance of high oleic acid in the seed oil of sunflower // Proc. of Sunflower Research Forum. - National Sunflower Association, Bismarck, ND, 1984. - P. 9.
- Urie A.L. Inheritance of high oleic acid in sunflower // Crop Science. - 1985. - Vol. 25. - No 6. - P. 986-989.
- Miller J.F., Zimmerman D.C., Vick B.A. Genetic control of high oleic acid content in sunflower oil // Crop Science. - 1987. - Vol. 27. - P. 923-926.
- Fernandez-Martinez J.М., Jimenez A., Dominguez J. Genetic analysis of the high oleic acid content in cultivated sunflower (Helianthus annuus L.) // Euphytica. - 1989. - Vol. 41. - P. 39-51.
- Perez-Vich B., Fernandez-Martinez J.М., Grondona M., Knapp S.J., Berry S.T. Stearoyl-ACP and oleoyl-PC desaturase genes cosegregate with quantitative trait loci underlying high stearic and high oleic acid mutant phenotypes in sunflower // Theoretical and Applied Genetics. - 2002. - Vol. 104. - P. 338-349.
- Velasco L., Perez-Vich B., Fernandez- Martinez J.М. Inheritance of oleic acid content under controlled environment // Proc. of 15th International Sunflower Conference, France, Toulouse, June 12-15, 2000. - Vol. 1. - P. A31- A36.
- Demurin Y., Škorić D. Unstable expression of Ol gene for high oleic acid content in sun-flower seeds // Proc. of 14th International Sunflower Conference, China, Shenyang, Beijing, June 12-20, 1996. - Р. 145-150.
- Demurin Y., Efimenko S., Borisenko O. A screening for suppressor genotypes on a high oleic mutation in sunflower // Proc. of 16th International Sunflower Conference, Fargo, ND, USA, August 29 - September 2, 2004. - Vol. 2. - Р. 779-782.
- Демурин Я.Н., Борисенко О.М. Устойчивость мутации высокоолеиновости масла к действию супрессора в семенах подсолнечника // Масличные культуры. Науч.-тех. бюл. ВНИИМК. - 2009. - Вып. 1 (140). - С. 18-21.
- Lacombe S., Abbott A.G., Berville A. Repeats of an oleate desaturase region cause silenc-ing of the normal gene explaining the high oleic Рervenets sunflower mutant // Helia. - 2002. - Vol. 25. - No 36. - P. 95-104.
- Hongtrakul V., Slabaugh M.B., Knapp S. A seed specific Δ-12 oleate desaturase gene is duplicated, rearranged and weakly expressed in high oleic acid sunflower lines // Crop Science. - 1998. - Vol. 38. - P. 1245-1249.
- Premnath A., Narayana M., Ramakrishnan C., Kuppusamy S., Chockalingam V. Mapping quantitative trait loci controlling oil content, oleic acid and linoleic acid content in sunflower (Helianthus annuus L.) // Molecular Breeding. - 2016. - Vol. 36. - P. 106.
- Schuppert G.F., Tang S., Slabaugh M.B., Knapp S.J. The sunflower high-oleic mutant Ol carries variable tandem repeats of fad2-1, a seed-specific oleoyl-phosphatidyl choline desaturase // Molecular Breeding. - 2006. - Vol. 17. - P. 241-256.
- Tilak I.S., Kisan B. Evaluation of SSR and INDEL markers associated with high and low oleic acid content in sunflower (Helianthus annuus L.) genotypes // Journal of Pharmacognosy and Phytochemistry. - 2017. - Vol. 6 (5). - P. 1560-1563.
- Dimitrijević A., Imerovski I., Miladinović D., Cvejić S., Jocić S., Zeremski T., Sakač Z. Oleic acid variation and marker-assisted detec-tion of Pervenets mutation in high- and low-oleic sunflower cross // Crop Breeding and Applied Biotechnology. - 2017. - Vol. 17. - P. 235-241.
- Miller J.F., Vick B.A. Registration of four mid-range oleic acid sunflower genetic stocks // Crop Science. - 2002. - Vol. 42. - No 3. - P. 994-994.
- Чебанова Ю.В., Борисенко О.М. Гетерогенность среднеолеиновых линий подсолнечника НА421, НА422 и НА424 по содержанию олеиновой кислоты в масле семян // IX всероссийская конференция молодых ученых и специалистов "Актуальные вопросы биологии, селекции, технологии возделывания и переработки масличных и других технических культур". - Краснодар, 2017. - С. 157-160.
- Demurin Ya., Skoric D., Veresbaranji I., Jocic S. Inheritance of increased oleic acid content in sunflower seed oil // Helia. - 2000. - V. 23. - No 32. - P. 87-92.
- Демурин Я.Н., Борисенко О.М., Чебанова Ю.В., Левуцкая А.Н. Материнский эффект в наследовании признака среднеолеиновости масла в семенах подсолнечника у гибридов первого поколения // Масличные культуры. Науч.-тех. бюл. ВНИИМК. - 2016. - Вып. 1 (165). - С. 16-21.
- Демурин Я.Н., Борисенко О.М., Чебанова Ю.В. Наследование признака среднеолеиновости масла в семенах подсолнечника у гибридов второго и третьего поколений // Масличные культуры. Науч.-тех. бюл. ВНИ-ИМК. - 2018. - Вып. 3 (175). - С. 3-8.
- Fernandez-Martinez J.M., Perez-Vich B., Velasco L. Sunflower // In: Oil Crops, Handbook of Plant Breeding, V. 4 / Ed. J. Vollmann and I. Rajcan. - Springer, 2009. - P. 155-232.
- Мельникова Ю.В., Рубцова С.Н., Пахомова Т.Н., Панченко В.В. Экономико- математическое моделирование конъюнктуры российского рынка высокоолеинового подсолнечника // Вестник Алтайской академии экономики и права. - 2019. - № 2-2. - С. 292-298.
- Литвиненко Г.Н., Терещенко Ю.А. Комплексный анализ производства и переработки высокоолеинового подсолнечника // Труды Кубанского государственного аграрного университета. - 2017. - № 64. - С. 31- 36.
- CODEX STANDARD FOR NAMED VEGETABLE OILS. Adopted 1999. Revisions 2001, 2003, 2009. Amendment 2005, 2011. - 16 р.
- Saghai-Maroof M.A., Soliman K.M., Jorgensen R.A., Allard R.W. Ribosomal DNA spacer-length polymorphisms in barley: Mendelian inheritance, chromosomal location, and population dynamics // PNAS USA. - 1984. - 81. - P. 8014-8018.
