The introduction of the domesticated horse in Southwest Asia

Author: Shev E.T.
Journal: Archaeology, Ethnology & Anthropology of Eurasia @journal-aeae-en
Section: The metal ages and medieval period
Article in issue: 1 т.44, 2016.
Free access
Short address: https://sciup.org/145145248
IDR: 145145248 | DOI: 10.17746/1563-0110.2016.44.1.123-136
Text of the review article The introduction of the domesticated horse in Southwest Asia
The domestication of the horse led to a dramatic reconfiguration of long-distance trade and warfare throughout Eurasia. There is a significant chronological gap between the earliest archaeological evidence of horse domestication at around 3500–3000 BC in the Eurasian steppe site of Botai in Kazakhstan and the subsequent adoption of the animal by Bronze Age civilizations in the Near East. During the Second Intermediate period beginning in the seventeenth century BC, the domesticated horse Equus caballus first appears in Upper Egypt, discovered in situ underneath a destruction layer dating to 1675 BC within the southern fortress of Buhen (Emery, 1960; Clutton-Brock, 1974).
A review of archaeozoological remains distributed throughout Southwest Asia has been conducted in this paper in order to establish a chronology of E. caballus dissemination and determine the avenues of domesticated horse dispersal from the Eurasian steppe into the Near East. To complicate the picture, the faunal record suggests the continual survival of the wild predecessor of domesticated horses Equus ferus throughout the Holocene in the southern Levant, northwestern Iran, and central Anatolia, areas in which it was previously thought to have become extinct.
Botai in Kazakhstan yields the oldest well-supported evidence of early horse domestication. Several lines of evidence suggest the inhabitants of Botai were keeping horses for consumption and for ritual purposes (Levine,
Equid remains from sites in Southwest Asia
|
ф о о О |
со |
.2 0 8 3 го о v> е 8 |
СЧ СО о> of О w со У ш. |
04 04 О О 04 С* о л < |
3 со го ф с го го го СП |
S S ГО е Ч |
Ю О о 04 ГО ф ф 1 |
о о сч ф ф Л го О |
о о сч _ф о < |
сч со сч о о о сч _ф о < |
со о о сч о п ф го с о Л ф |
с |
ю со 7 со со со о о сч о Л ф го ^ |
|
т о "ф о с СП го Q |
< Z |
Е Е со л Л го го о 5 Е £ CL |
< Z |
Л о о "с го ф ф "5 < о- A s ш Z |
о со О — А о О г Li • & 5 5? о -S.CO й ^ ” ® ” 5 О. L К СО " см со со oS g 5 2 s s Б s8 S Sg “ ё о о о 3 с о^2сосм + см® < 2 CL 2 CL CL CL CL 2 |
< Z |
ф ф с го о го о |
||||||
|
s 2 »■ С З С 8 5'6 £” б |
со |
< Z |
со |
< 2 00 z £ сч |
z S S |
ю со ^ сч ю |
< 8 . 8 8 8 8 8 8 Z |
ю 04 о |
< Z |
||||
|
CL О Z |
ю |
< Z |
8 |
8 |
^ со о сч ^ ^ |
ю -<- ^ СЧ ^ |
< 8 8 Z |
т- Т- Т- т- т- т- т- т- т- |
|||||
|
т Ф 0) О |
"Г |
V) ГО «о ГО о ui |
6 |
ф" = = ui |
ф го «о го 8 8 8 О ui |
Uj |
V) V) V) го ro.ro -Q -Q CL -Q ГО SS 8 ГО 8 CL ГО 8 О О Ф О uj uj uj uj |
6 |
V) го -Q ГО О ui |
||||
|
с о го о —1 |
со |
< Z |
Е 3 см > ф Ф _ о ф 1 О-СМ $ = W % -1 И см" ^ см" > Ю ^ 0) СП ф СП —1 .Е р го р с го О "5 го 00 03 СП |
ф ф < < = Ф Ф 03 с "с ф О О ±± О О (У) |
о" со ш 2 > ГО ГО (V Ф ф ф > > ч- ф ф < —1—1 |
о О ОО 0 0^00 О ОО О СО О О О) со О СО О О СО СО СО СО Н О ОО ^ О О 2 ° ° СО ^ со СЧ Ю СО - О Ю о h- о со ю ю со +- г- < о + —1 —1—1 —1 —1 —1 —1 —1 —1 |
< Z |
о ф о —1 X ш ф го о |
|||||
|
ф СП < |
СЧ |
о СП Е "Е ф ^ О —1 |
о СП о о ю со 1 о о о О —1 |
о ш со со со со со СП ш |
О о Ю о л = о 8 СО zg Ф г-ГО ю —1 |
о о +1 +1 о о ОО „О со со О ГП сч сч £ о -I- -I' ” ° ° z о +1+1 ст ш со С оо ГО ^ т- т —1 ю ю |
О О 0Q СО ГО го о о ..О -О ОО ОО • F о ' F о со хГ Я О go £ о -Е о го сч го сч И СО ЛЮ О О |
О ОО 00 . СО 0Q О о 2 О О 03 2 О 2 8 § 7 2 7 7 е? о со о о Д о -о о 2 ю Q2 Ю со 2 СО Ш 00 ^2 О О О CQ <( |
о ш о о со 7 о о о сч СП |
6 7 ° ^ £ сч = сч О ЧТ ф 1 Z О £ со го ю _। |
|||
|
ф О |
th Ф Ф ф Е ф 5 |
го о |
Е о о т |
о Л ГО го О |
ГО ГО с п_ |
о т ■фп о |
о т го О' |
го фл ф |
с "го р N ГО О л" ф л СП го N |
|
LO 7 o> o> Ф о |
ю со 7 со со со о о СЧ о tn 05 ^ |
я СП |
o' сч со" со о о сч го" |
со со со С Ф с О .с о ел я ^ |
е S ф £ 3 о V) |
СО О сч с го В со со 1 ■Q 03 ш ” |
со LO СЧ ю о о сч ф |
о о LO 05 О) с" О ел го о £ |
О) о °? ш^ |
ф ^ ел -О .£ к N" g g S' ° щ |
О о сч N 1 о т |
со О) О) с о tn О) о |
|||
|
i ® S 05 CO" CO CL |
” Й = S E" 2 X ГО , me Ф О ф 3 го ф E Я E Eg Я „ E П Ji aS - CD co I 2 W IL H 2 CL W |
ф" ф С 05 ^ о ^Г 05 О ел tn ^ ф Е Е Е т |
- 8 7' tn ^ го ел ел Ф го с Ф Ф О ГО £ ГО — ГО О 05 Т О |
tn ф £ П ГО Ф —" ГО Я & 5 03 |
сч т § п о о го "5 а? го со |
< |
ел 'S- ГО ГО р ГО Ф ф — Е 5 'х i5 (X |
ел Ф О) с го го |
ф го о го ф ^ |
го о го ф Е го "ф b |
го о го ф Е ф .5 § 2 т |
8 ^ Я 5 ” Ф Го -О ^ Z н |
|||
|
5$ 8 Z |
< г |
= |
7 |
< |
^ со о о |
V |
V |
V |
V V V |
||||||
|
--- CO |
. 8 . . € € Z м Z CN CN |
со |
8 |
со |
ю |
сч -<- |
со |
со |
|||||||
|
ел СП «о СП о ui |
я ui |
ел (П -Q ГО s О ui |
ел g ui |
ел § (П «о го о ui |
ел го -Q ГО О ui |
6 6 6 |
|||||||||
|
X Ф § 3 |
ф z — - Z z z = z - ^ Ф > ~ ^ Ф i_ Ф io m z cd io io io $ J -1 $, m J -> ro -> in co co -1 co" in . co" J ® К -1 ^ O> CO CM CD M" cm t- см см т cm Б £ < 5 5 £ § ^ § c c c c c c c c c ZD ZD ZD ZD ZD ZD ZD ZD ZD |
со о ^8 ф з го о ГО о СО |
сч izE со" со" X X X X о о 5 £ Е ф 05 го >> ГО ГО ГО СО СО |
о о о ф ел Ф £ |
< г |
< г |
СП ф го —1 |
< о ф ф (У) (У) |
< г |
О) сч ф 1 |
со о сч СП сч" о сч —1 ф го о |
со со со Ё Е (У) |
О со О 05 05 05 Ф Ф Ф < < < |
||
|
о Ш o 8 Ш О 8 8 ш О
1 1 § A S --8 Б § з
- —" = = о — — — — со со со со со см ш ш ш ш |
о LO +1 о со 7 ■I-о LO +1 о - 05 СП сч ш |
о со +1 о со ■1- о сч । со СП ^“ —1 |
О СП о ю со сч 05 О СП ш |
о СП о ю LO 7 о о сч 05 О СП |
о СП о о о сч" „ 7 с о 05 О ч= Ю 5 сч" 05 т- г |
о СП го о о со о" - 7 с со ■- § го 7 г |
о (И о. о - о rQ О CD CD ш О z ю Z со ю CL О Ll о ф ю Го о Го со . С СО |
о СП о о „ о С LO у О е ° О ю Е ГО ^ О |
Я" О £ со ° 8 8 у ГО 05 С> —1 |
S го ф го о о го о о —1 |
о (П о о со со о о со со о —1 |
ф Е 05 (У) |
|||
|
0) £ с о О |
с "го 0_ n 05 а с ГО 'tn ф го а |
с "го 0_ с N 05 а ГО ГО N О) ГО СО |
го ф -С О |
го го го го т |
сч JZ ф Е £ 05 С Т 05 р |
ф го ел ф" го О Е "с о го т |
с 05 О С 05 С "ф I |
с го о 05 DZ < |
Ф ГО ф Е "с о го 05 (У) ГО Е 05 DZ |
ф го ф о с Е ГО п о (У) |
ф го ф Е Е (У) |
Ф ГО ел го о |
Table (end)
|
co |
o' o' cd co b- CO ь- Ob- о ^ О ч- ГО co" ГО E > -6 E 5 Q S |
о о СЧ го со го ^ |
СЧ 3 СЧ О О о СЧ СО = S' |
СО о о СЧ ГО ГО < |
о со о> О) го - 0? О) *у-с CN S ci £й |
с о со ю со го 2: СП со с . . > со Го ^ т |
0? о с го О |
со о> ю 3 ю c\i о CN го —1^ |
o' LO О сч с го |
ю о> о> с о го о ^ го о 5 О |
со о о сч го" о о |
|
CD О CN CO О с ь. co c ГО (D M S Q) c 00 55 ю ГО ГО т to ГО CL X Q. X c x С 05 ГО c 5 ф ■§. ф 5 CL 2 2 CL |
~ го го S го 5 "го "ю h h |
го о = 5 |
< Z 8 8 8 |
& ^1$ g “ “ X см 2 2 с 5 с го СП 5 О О < s”5 X ® а х - |
< |
го ГО СП го "го < |
со 2 см" 2 Z |
< Z |
"с го го го О . о го .41 = о X Го ГО С |
< Z |
|
|
co |
< 8 z |
8 8 |
ь- ^ < |< о z |
СО со ^ ю о Ю CN ^ ^ |
О) ^ М т о со о о о |
< г |
< Z |
8 |
8 |
8 |
со о |
|
LO |
т- 04 т- ч- |
о ^ СО ^ |
£ 8 £ S ю ю ^ ь- |
-С- CN OJ -с- ю |
S со |
8 со |
S о |
||||
|
"Г |
to to to tU tU tu -Q -Q -Q ГО ГО a ГО О О о Hi uj uj |
to СП ГО 8 8 о ui |
6 6 6 |
= 8 = 8 |
6 |
ГО СП «о СП о ui |
6 |
||||
|
co |
co -<- о ^ 04 co co > о о — — — _| = _| = — н- ф i— ф | 1 2 5 2 5 5 со < < со |
и_ О го го ГО 0) < < |
СП < ГО 8 ГО ГО ГО < < |
ГО -Q СО ГО СО СО СЧ СЧ |
"(О ГО 8 8 8 8 го < |
о со го Q o' го го < |
CN о о 9 го ГО о =5 О о со СП < |
Ч-Г 00 § Ё оз -5 2 55 < |
1 |
Е го со |
|
|
> > > > |
|||||||||||
|
О о о о < < < < |
|||||||||||
|
СЧ |
о Ц О о СО 00 ю ° о 5 +| 78 К 88 Si S3 ^ ° ° 04 “ о со -со - О — = СЧ — 04 — — СЧ СП СО СП СП СП СЧ ш ш ш ш |
о ап о о о со о о со со СП ш |
о О S “ 8 О ь~ ю ^ СО 1 СЧ О 1 О о о Ю СЧ СО < сп оп ш ^ |
о о со CQ о о 2 ° ч Ч ci о ° О 0x1 СЧ СП СП ш ш |
о о СП “ о 1 § Q ш О 1 § § О ” ® L см g со со го 7 ч 8 о СО СО g СП с = "L ° ё о СО пп CD СО CD 2 _l S CL I |
О ап о ю со 7 о о LO о LO ю ап —1 |
О ш о ю 7 о о ю ап —1 |
о ап о о о 7 о о со < |
О ап о о о 7 о о со < го го —1 |
о ап о со ю о сч сч < |
о ап со со сч со о "го "с го го т |
|
ГО го со го < |
ГО го со го 5 |
го го со го < 5 |
го го СП ^ с .!= О 5 |
го го со го о |
го го со .с о ^Е со |
го го го го го со |
го го со го О го со го го |
го го го с го Q |
с го о с го |
го го го Е" го £ го ап |
Note : LC – Late Chalcolithic, EB – Early Bronze Age, MB – Middle Bronze Age, LB – Late Bronze Age, IA – Iron Age.
*In parentheses, MNI is given (if known from the source).
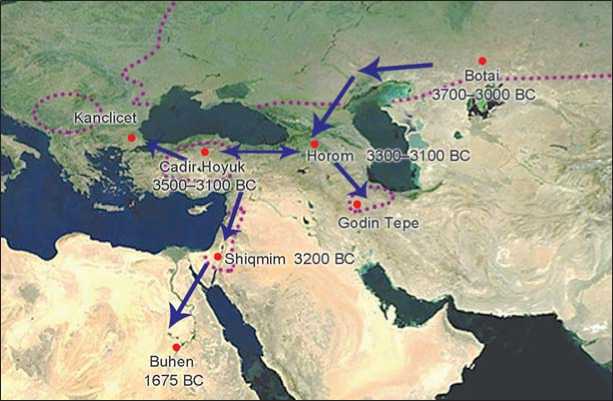
Fig. 1 . Possible dissemination of E. caballus in the Near East, and key sites discussed in this paper. Dotted lines indicate regions were wild horses continued to exist during the midHolocene (ca 5000 BC). Dates indicate the earliest evidence of domesticates within certain regions.
1999; Olsen, 2003: 98–101). Analysis by Brown and Anthony (1998) yielded at least four individual horses that demonstrate evidence of bit-wear on horse lower second premolars (P2), a pathological indicator of domestic status.
Interestingly, the dissemination of the domesticated horse has been linked to the spread of Indo-European languages. For this reason, archaeozoological evidence indicating the geographical spread and chronology of horse dissemination has long been entangled with the Indo-European question. Gordon Childe (1926) highlighted commonly shared linguistic elements, or cognates, within ancient and contemporary IndoEuropean languages in theorizing of a common ProtoIndo-European culture. The Proto-Indo-European link to horsemanship is one that may have some credence given that the first domestication event likely took place in roughly the same region that was the homeland of all Indo-European languages.
The transmission of the domesticated horse from the Eurasian steppe into the Near East could have only occurred via a limited number of geographical corridors. As it stands, there is no evidence that horses were present in the Balkans or eastern Iran at a period contemporary with or shortly after horse domestication occurred at Botai. Transcaucasia, Anatolia, and northwestern Iran on the other hand do demonstrate populations of E. ferus/ caballus between 3500–3000 BC, indicating Transcaucasia as a likely corridor of dissemination (Fig. 1).
Archaeozoological method for identifying Equus caballus
To ascertain the presence of equine species in the faunal record, an understanding of archaeozoological methods of identification is needed. All equid species share similar post-cranial elements so the accurate identification of species can be problematic. Determining domestic status is another problem; early examples of domesticated horses do not exhibit the clear phenotypic variations from wild species that are observable in other domesticated animals such as the bovids. Aside from osteological traits, domestic status can be determined by diagnostic pathological traits such as bitting scars, and also from observable changes in butchery practices and the presence of artifacts such as riding equipment and objects that exhibit the horse as a ritual symbol (Anthony, 2007: 216– 218, 221, 239).
Teeth are the most frequently encountered skeletal remains and are often used to distinguish between E. asinus/ hemionus , E. hydruntinus , and E. ferus/ caballus (Davis, 1980; Vila, 2006: 103). Observations of the buccal (external) fold and its depth of penetration towards the lingual (internal) fold is one method of identifying species. Caballine molar lingual folds are U-shaped and are the most recognizable dental feature attributable to horses (Davis, 1980; McGrew, 1944).
Distinguishing between wild and domesticated status from caballine remains is also problematic. Unlike the case with other domestic animals such as bovids, variations in body size are not a reliable indication of domestic status in horses. Shifts in size may be related to the transmigratory habits of horse populations rather than due to controlled breeding (Anthony, Brown, 2011). On the other hand, certain elements of E. caballus skeletal morphology can exhibit alteration from that of wild stock; metapodials can undergo morphological change due to an increase in their load-bearing function when horses are utilized as beasts of burden (Outram et al., 2009). Wear on dentition is also another observable trait that has been linked to human actions. The bitting of horses to fix harnesses can create scarification and wear on mandibles and teeth, characteristics that are pathological indicators of domestic status (Anthony, Brown, 2011). “Type 1” bit- wear is caused by the pulling of reins and consists of a beveled edge of more than 3 mm on the occlusal surface of the mesial corner of P2 molars (Anthony, Brown, 1991; Brown, Anthony, 1998).
Regions examined
To establish the chronology detailing how the domesticated horse came into use in Mesopotamia and Egypt during the late 3rd and early 2nd millennium BC, an understanding of the faunal record of neighboring regions that are probable geographic routes of dissemination is needed. Between the Eurasian steppe and the Levant, Iran and Anatolia, lies the Transcaucasia region.
Transcaucasia. Domesticated horses likely disseminated from the Eurasian steppe into the Near East via Transcaucasia (Fig. 2). Horses appear in the Caucasus during the Maikop and Early Transcaucasian (ETC) cultural periods between 3500–3000 BC (Anthony, 2007: 221). Given that there is no evidence of wild early Holocene populations of horses throughout the Caucasus, they were likely imported either through the Pontic-Caspian steppe or via eastern Anatolia.
One of the earliest instances of E. caballus within the region dates to the late 4th millennium BC from the site of Alikemektepesi, Azerbaijan, in the steppe south of the Kura and Araxes rivers (Mallory, 1989: 232). At Alikemektepesi, two species of equids were identified, the larger of which was thought by Soviet archaeologists to be the earliest evidence of domesticated horses in the Transcaucasian region (Eneolit SSSR, 1982: 134– 135). The dating of these specimens suggests an early intrusion of horses from the Pontic-Caspian steppe and is synchronous with findings in Anatolia (Grigoriev, 2002: 336; Mallory, 1989: 232–233).
A more secure dating of what may be a domesticated horse comes from the site of Mokhrablur, Armenia, and dates to the Late Chalcolithic. Evidence of domestication is derived from bit-wear analysis on a 5-year old stallion discovered at Mokhrablur. Severe wear on the mesial edge of a P2 molar demonstrates a bevel deeper that 3 mm, suggesting that this animal was bitted (Anthony, 2007: 221). The presence of this specimen in LC layers suggests that there was a dispersal of E. caballus into Transcaucasia prior to 3000 BC (Anthony, Brown, 1989: 102).
The site of Horom is located within the Shirak Plain in northwest Armenia and boasts an occupational history spanning around 5000 years, ranging from the EB I– Early Iron Age (IA I). Radiocarbon sampling of EB I material (sample no. AA-7767 at 1 sigma) taken from the 1990 sounding of the site gave a date range of 3371– 3136 BC (Badjalan et al., 1992, 1993). During the 1992– 1993 seasons, over six thousand faunal specimens were uncovered, many of them well-preserved. The domestic assemblage accounted for 92.4 % of all identifiable remains, suggesting an agro-pastoral subsistence strategy. Horse bones comprised 6.7 % of the EB I–IA I assemblage (Badjalan et al., 1994).
Anatolia. Recently uncovered evidence suggests the continued survival of E. ferus in central and eastern
Qadir Hdyuk
Horom
Pinarba^i
Kosk Ho;
Qatalhoyuk
Tell Habuba Kabira
Godin Tepe
♦а e b
Hayonim Cave
Tel Aphek \
Tel Dor. у
Tel Michal X
Shoham у'
?k Afridar У
Qabrestarf
Zagheht* Sagzabad
Fei Dan
*Ain Rahub
Wadi Hammeh 27
>УТе1 Rehov x Khirbet al-Baltrawy
Shiqmim"-^yad 5hiloh , . ^ ^/vadi Feinan lat Saharonim •petra
•c •e •g i df•h j
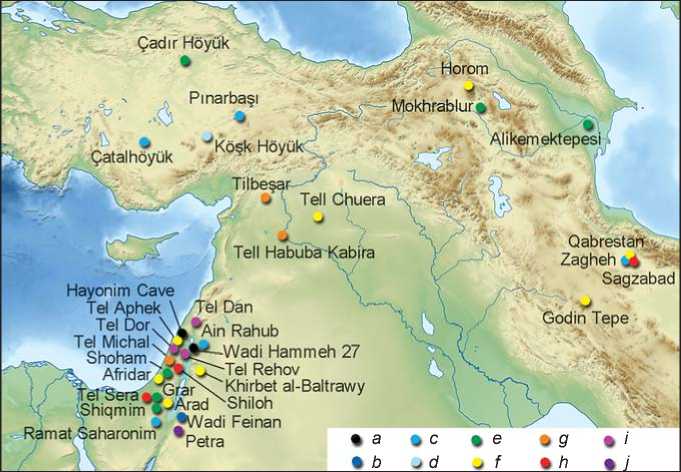
Fig. 2. Chronology and distribution of E. caballus remains in Southwest Asia.
a – Natufian; b – PPNB; c – Late Neolithic; d – Early Chalcolithic; e – Late Chalcolithic; f – Early Bronze; g – Middle Bronze; h – Late Bronze; i – Iron Age I; j – Iron Age II.
Anatolia well into the Holocene, confounding attempts to draw a linear direction of dissemination from the Eurasian steppe into the Near East. It is widely accepted that horses became extinct throughout much of the Old World at the end of the Pleistocene, existing only in scattered pockets throughout Europe (Anthony, 2007: 198). Contrary to this, recent research has indicated that wild horses were present throughout central and eastern Anatolia well into the Holocene. Faunal assemblages from the Neolithic settlements of ^atalhoyuk and Pinarba$i indicate that a small percentage of wild horses were hunted in the central plateau between 7400–6200 BC (Martin, 2013; Carruthers, 2013). Importantly, E. ferus remains have been uncovered across the central plateau during the 4th-3rd millennium BC, both in the Altinova Plain in eastern Turkey and the Eskisehir region in the northwest of Anatolia (Arbuckle, 2009).
The Cappadocian region site of Ko5k Hoyuk demonstrates a prevalence of the consumption of horsemeat dating to the 6th millennium cal BC. Five distinct occupational levels have been determined at Ko$k Hoyuk encompassing the Neolithic to Chalcolithic periods (Arbuckle, 2007). The majority of equid remains were uncovered in areas E–F/8–9 from various pit features located on the fringes of the settlement, designated for the butchering of large animals (Öztan et al., 2005). Lower Phases V–II (6200–5200 cal BC) contained substantial quantities of equid remains, accounting for 23 % of the total assemblage. In some contexts, equid bones accounted for 50 % of the faunal remains. Of the three equid species uncovered at Ko$k Hoyuk, E. hemionus and E. hydruntinus were represented by smaller-sized remains, and larger bodied equids fell into the size range of E. ferus (Arbuckle, 2007; Martin, Russell, Carruthers, 2002).
The Chalcolithic site of ^adir Hoyuk raises the prospect of an independent area of domestication (Sagona, 2011: 696). ^adir is one of many settlements that appeared in the Kanak Su basin during the 4th millennium BC. The stratigraphic sequence of the mound demonstrates continuous occupation from the Middle Chalcolithic (5200 cal BC) to 1100 AD (Arbuckle, 2009; Steadman et al., 2008). There is an abundance of equid remains at the site, accounting for 3 % (NISP = 71) of the total assemblage for all periods. Equid remains fluctuate from 14 % in the Late Chalcolithic/ Early Bronze Age assemblage to <1 % during the Byzantine era. Equid specimen CD2015 from Late Chalcolithic (LC) deposits (Late Chalcolithic Phase Ib, locus 62, trench 770.900) illustrates the presence of horses from the first half of the 4th millennium cal BC. Five separate specimens from Iron Age (IA) and LC/ EB I contexts exhibit butchering marks on metatarsals, metacarpals, and the distal aspect of one radius. Equid remains with butchery marks account for 7 % of total assemblage, reflective of subsistence patterns illustrated in some Eurasian steppe sites (Arbuckle, 2009).
There is no observable biometric variation in specimens from the LC to the IA, suggesting a steady continuation of phylogenetic morphology. This would not be expected if domesticated horses from a different region had been introduced in the 4th–3rd millennium BC. Many specimens from the Late Chalcolithic exhibit a high proportion of foot bone pathologies; 20 % (NISP = 30) of the specimens show pathological degradation that may be the result of being utilized as beasts of burden (Ibid.). If this data can be relied upon, the central plateau of Anatolia may represent an independent area of horse domestication.
Northwestern Iran. Northwestern Iran lies on the southeastern border of the Transcaucasia region and is therefore a likely candidate for the dispersal of E. caballus southward at the end of the 4th millennium BC. There are a few sites that demonstrate the presence of domesticated horses, both in the Qazvin Plain and also in the mountainous Zagros region. However, there is also evidence that, like eastern Anatolia, western Iran may have also supported small, isolated populations of E. ferus well into the Holocene. Remains of E. ferus are present in small numbers within the Chalcolithic site of Zagheh, northwest of Tehran. These specimens precede the earliest evidence of domestication in the Eurasian steppe. Radiocarbon samples have established a site chronology demonstrating occupation from Late Neolithic/ Transitional Chalcolithic to the Early Chalcolithic ca. 5370/5070–4450/4220 cal BC (Fazeli, Wong, Potts, 2005). E. ferus skeletal elements at Zagheh include teeth, a humerus, a calcaneus, and several second and third phalanges dating from the 6th millennium BC (Mashkour, 2003: 134–135). These specimens represent a further geographical extension of Holocene era E. ferus than previously thought, just as recent research has revealed a Holocene presence of wild horses in central and eastern Anatolia.
Qabrestan in the Qazvin Plain demonstrates the presence of horses at a later date, contemporary with examples uncovered in central Anatolia and Transcaucasia. At Qabrestan, an equid calcaneus that fits into the size range of E. caballus was uncovered, dating to the early 3rd millennium BC (Ibid.: 134).
Godin Tepe is located 1600 m above sea level in the Kangavar Valley of western Iran. Its stratigraphic sequence contains at least ten defined cultural phases, the earliest of which dates to around 4500 BC (Young, 1969). Godin IV (3000–2600 BC) is marked by evidence of an intrusion by Yanik/ ETC culture from the north; it also contains the oldest E. caballus remains at the site (Burney, Lang, 1971: 52–53; Gilbert, 1991: 75–76; Young, 1969: 19–20). During the 1967–1973 seasons, excavation of Godin III and IV yielded a substantial assemblage of faunal material (Gilbert, 1979). Equid bones are best represented during Godin IV (NISP = 146), the majority of which were identified as E. hemionus. The remaining 6 % of the equid remains were determined to fit within E. caballus size ranges (Gilbert, 1991: 78, 87). Two securely identifiable caballine remains were uncovered from Godin IV/V, a complete right metacarpal III, and a right mandibular cheek-tooth. The metacarpal exhibited the caballine traits of mediolateral eminences and a weak dorsal relief (Ibid.: 96).
Determining species from the cheek teeth was problematic due to poor preservation conditions and the presence of hybridized traits. However, many of the analyzed cheek teeth also demonstrated the caballine trait of sulcus externus , a deeply penetrating metalophid fold between the hypoconid and protoconid cusps. Of all analyzed cheek teeth, only two individual specimens demonstrated solely hemione traits, while only one demonstrated solely caballine traits; even so, this single tooth also shows minor signs of blending (Ibid.: 88). Gilbert (pers. comm., 2013) is currently analyzing the DNA of faunal remains from Godin IV to examine the possibility of equid hybridization at the site.
Syria. Given the likely northern Eurasian steppe or possible Anatolian origins of the domesticated horse, it is logical to investigate the archaeozoological record of Late
Chalcolithic and Early Bronze Age sites in the northern Levant as a means of dispersal to the southern Levant and Egypt. Several species of equids existed in Southwest Asia during the Pleistocene, however increasing aridity occurring at the beginning of the Holocene period saw equid diversity diminish dramatically and a decrease in previously abundant E. ferus populations (Orlando et al., 2009). However, there are a number of settlements in southerly regions that demonstrate the continual presence of wild horses throughout the Holocene (Fig. 3, 4).
There are only scattered examples of 3rd millennium BC remains found, most significantly at the site of Tell Chuera in northeast Syria. Tell Chuera is situated between the Khabur and Balikh rivers approximately 11 km south of the Syrian-Turkish border and covers an area of around 65 ha. The settlement was possibly established by migrants from the wetter areas of the Upper Khabur in Anatolia at around 2900 BC (Akkermans, Schwartz, 2003: 256, 259). A number of equid bones were uncovered at Tell Chuera that exceed the size range of E. asinus and E. hemionus . Uncovered from the temple sector (Tell Chuera IC), dating to the middle of the 3rd millennium BC was a first phalanx, the distal extremity of a humerus, and a scapula, all of which were within caballine size ranges.
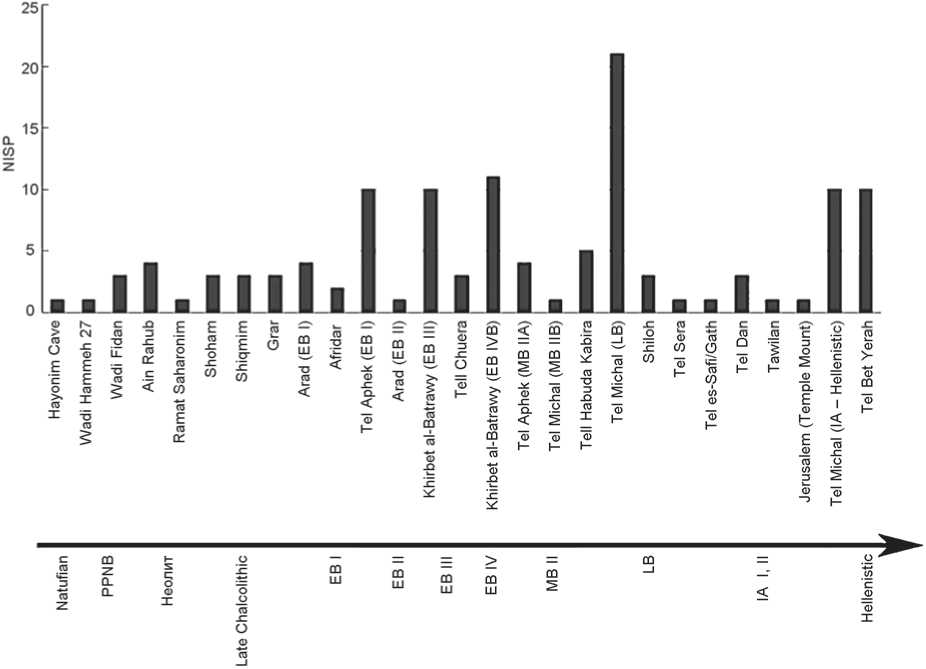
Fig. 3 . Number of individual specimens (NISP) of E. caballus remains from Levantine sites, ordered chronologically. PPNB – Pre-Pottery Neolithic B, EB – Early Bronze Age, MB – Middle Bronze Age, LB – Late Bronze Age, IA – Early Iron Age.
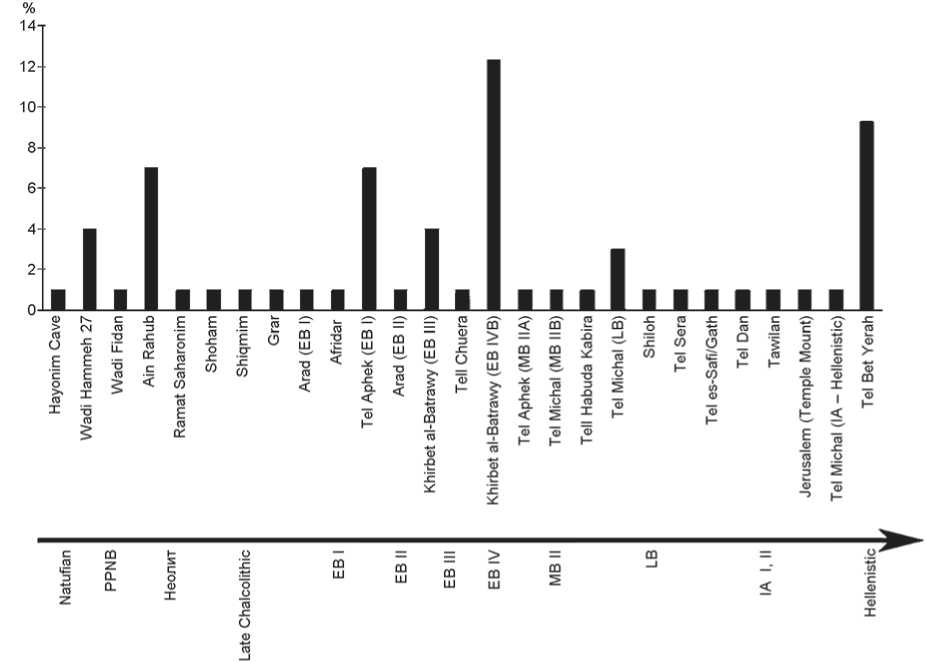
Fig. 4 . Percentage of E. caballus remains within the total faunal assemblages from Levantine sites, ordered chronologically.
Legend same as on Fig. 3.
These specimens indicate the presence of domesticated horses in Syria at around 2650 BC (Vila, 2006: 118, 120).
Southern Levant. Wadi Hammeh 27 in the Jordan Valley is a Natufian period settlement that contains evidence of E. ferus in the southern Levant at the end of the Pleistocene. Wadi Hammeh 27 dates to around 12,000 cal BC and contains early evidence of the harvesting of wild grain (Edwards, 2007). Faunal analysis revealed that equids account for 2 % of the total NISP at Wadi Hammeh 27. A single mandibular tooth (specimen WH27-832) was classified as belonging to E. ferus due to its U-shaped lingual fold with an intermediate penetration towards the lingual fold (Edwards, Martin, 2013: 337). Further evidence of the presence of E. ferus during the Natufian period is exhibited at Hayonim Cave in Upper Galilee. Contained within Hayonim Layer B (11,403– 10,370 cal BC) was found a single post-cranial element that was identified as E. ferus (Stiner, 2005: 256).
Remains of E. ferus from the ceramic Yarmoukian period are found in northwestern Jordan. The site of ‘Ain Rahub is located on the lower terrace of the west bank of Wadi er-Rahub, approximately 13 km northeast of the modern settlement of Irbid (El-Shiyab, 1997: 593). The later Yarmoukian layers however revealed four first phalanges identified as horses, accounting for 7 % of the total identifiable faunal assemblage of this period. These phalanges were identified as E. ferus due to their solid middle shafts and the presence of developed distal supra-articular tuberosities (Ibid.: 594– 596). Contemporarily with the Yarmoukian specimens of ‘Ain Rahub is the presence of an equid metacarpal that falls into the size range of a small horse found at the site of Ramat Saharonim in the Negev Desert (Horwitz, Rosen, Bocquentin, 2011). Excavation of Tumulus 28–30 and Shrine 4 fixed a contemporaneous date of around 5000 BC for the structures (Porat et al., 2006; Rosen et al., 2007). Discovered in context with human remains was a single, fused distal metacarpus III, osteometric analysis of which affirmed a size within that of the E. caballus range, similar to the Late Bronze Age Egyptian Soleb horse (Clutton-Brock, 1974). According to Horwitz, Rosen, and Bocquentin (2011), this specimen may be representative of either a large previously unidentified equid species, an unusually large E. africanus, or the continued habitation of the southern Levant by E. ferus.
The Late Chalcolithic burial caves of Shoham (north) in the Lod Valley may contain evidence of the presence of E. ferus. The faunal assemblage of Cave 4 demonstrates an abundance of equid remains accounting for 32.8 % of the total identifiable skeletal elements. Of these, 226 specimens were identified as belonging to E. asinus, while three postcranial elements were acknowledged as belonging to a larger equid species, either E. caballus or E. africanus. A single distal metapodial derived from context L202, B2043, exhibited a breath measurement (Bd) of 47.9 mm (Horwitz, 2007). The mean breadth measurement for E. caballus distal metapodials is 52.3 mm, indicating that this specimen could be within the size range of a horse (Clutton-Brock, Burleigh, 1979). However it must be noted that these three postcranial elements identified as large equids were discovered within the baulk just below the topsoil, rendering their dating as ambiguous.
Several sites in the northern Negev demonstrate equid remains that fall within caballine size ranges. One of these sites, Shiqmim, contains a rich assemblage of cultural material and evidence of relatively advanced metallurgy dating between 4500–3200 BC (Gilead, 1989). During the 1989 excavation season at Shiqmim, the proximal humerus of an equid was uncovered that was comparable in size to specimens of E. caballus from the Roman period site of Gamla (Grigson, 1993). The Shiqmim humerus (bone no. shq 89.1054) had a proximal breadth of 92.6 mm, compared to the Roman period specimen from Gamla at 91.2 mm (Levy et al., 1991). The prospect of intrusion from later strata seems unlikely given that seven horse bones were uncovered from well-stratified Chalcolithic layers at two different Negev sites, Shiqmim and Grar. The Shiqmim humerus was discovered above the floor of an underground room that had been sealed by aeolian fill radiocarbon dated to 3240 ± 75 cal BC, thus confirming a 4th millennium date for the specimen. Additionally, metacarpals attributed to E. caballus at Shiqmim demonstrate exostoses on the sides of the distal end of the shaft, indicative of osteopathic wear associated with draft-labor (Grigson, 1993).
The Early Bronze Age city of Arad in the northern Negev contains the remains of a large equid dating to a few hundred years after Shiqmim. Arad is situated at an elevation of 550 m, approximately 31 km east of the modern day city of Beersheba. The city contains occupation levels dating to EB I and EB II within strata IV–I, roughly corresponding the First and Second Dynasties of Egypt, 3100–2950 BC and 2950–2650 BC respectively (Davis, 1976; Kuhrt, 1995: 118). The faunal assemblage from the 1971–1974 seasons contained a single metacarpus that falls into the size range of E. caballus. Several equid remains were uncovered at Arad including numerous phalanges, three intact metacarpals, and one left mandible containing cheek teeth. Of the three metacarpals, one specimen was comparatively robust (Davis, 1976). Osteometric analysis of the robust metacarpus (specimen no. 8672, locus 4114, layer II) yielded a maximum length of 219 mm and a proximal width of 51.8 mm. Comparing this to a known domesticated specimen from Soleb (Egypt, Dynasty XVIII) indicated that the Arad bone was notably larger. The Soleb specimen measured a maximum length of 215 mm and a proximal width of 48.6 mm, indicating that the Arad specimen fell into the range of domesticated horses (Clutton-Brock, 1974; Davis, 1976).
Salvage excavations conducted in the Afridar neighborhood of Ashqelon have revealed an extensive EB I settlement (Golani, 2008). Four thousand identifiable animal bones were uncovered at Afridar, making it one of the largest EB faunal assemblages in the southern Levant. Most of the diagnostic equid remains were identified as E. asinus , verifying an early spread of domesticated donkeys into the region from Egypt, the earliest evidence of which was found at El Omari dating to 4600–4400 BC (Boessneck, Driesch, von den, 1990: 99–101). There were a few post-cranial elements that exceeded the size range of E. asinus and corresponded with that of larger equids . Area E, Locus 33 contained thirty-two equid bones, mandibles and maxillae, comprising the majority of remains (NISP = 27). The assemblage represents an MNI of four individual equids, three donkeys, and one horse. Contained within Areas F and G were two large equid limb bones, a distal radius with a breadth of 66.4 mm, and a distal tibia with a breadth of 74.5 mm (Kansa, 2004). These measurements fit well within caballine dimensions, suggesting that wild horses persisted in the southern Levant, if indeed these specimens were not domesticated.
The appearance of E. caballus in Egypt by the 17th century BC is best explained by the transmigration of technology and trade from the Levant to the northeast. There are three Early to Middle Bronze Age sites in the southern Levant that contain E. caballus remains that are largely contemporary with or closely predate the Buhen horse: Tel Aphek, Khirbet al-Batrawy, and Tel Michal.
Due to its strategic location adjacent to the headwaters of the Yarkon River and its situation along ancient trading routes, Tel Aphek contains an extensive history of settlement spanning several occupation events from Early Bronze I Phase C (3150–2850 BC) until the early 20th century AD. Tel Aphek was one of a number of fortified settlements that emerged in the Levant at the beginning of the 3rd millennium BC. Contained within the settlement are two areas that contained remains of E. caballus/ ferus ; areas A and B (Kochavi, 1977: 15). Within Early Bronze I strata of Area B were found ten E. caballus/ ferus skeletal fragments, accounting for an MNI of one individual horse. Within Middle Bronze IIA strata of Area A was uncovered one fragment within prepalace Area A and three fragments within Area B strata, these remains dating to approximately 2000–1750 BC. The majority of identified E. caballus remains consisted of teeth (Hellwing, 2000: 294, 297, 305).
The Jordanian Early Bronze Age site of Khirbet al-Batrawy, excavated since 2005 by La Sapienza University, clearly demonstrates the presence of E. caballus/ ferus from the EB III during the mid-3rd millennium BC. An earthquake around 2700 BC destroyed much of the site however the settlement subsequently increased in size after the disaster (Nigro, 2013). Within Activity 3a and 3b EB III layers dating after the earthquake, at least ten caballine skeletal elements (MNI = 3) were uncovered. A further eleven caballine bones were discovered in EB IVB layers, indicating the presence of E. caballus at the site from around 2700–2100 BC (Alhaique, 2008).
E. caballus remains chronologically closer to the dating of the Egyptian Buhen horse were found at the settlement of Tel Michal, located on the coastal plain of Israel approximately 12.5 km north of Jaffa (Herzog, Rapp, Muhly, 1989: 3). Tel Michal was originally settled during the Middle Bronze Age IIB (1800–1550 BC) and was occupied until the early Arab period. Site A, covering an area of approximately 1600 m2, contains the majority of studied cultural and faunal remains (Herzog, Negbi, Moshkowitz, 1978; Herzog, Rapp, Muhly, 1989: 5).
Faunal analysis of Middle Bronze Age strata revealed small numbers of E. caballus remains, accounting for 0.9 % of the total assemblage. Late Bronze Age (1700–1200 BC) strata revealed substantial amounts of E. caballus remains at 21 fragments (MNI = 1). Within Iron Age, Persian and Hellenistic era strata E. caballus remains were also uncovered, however it must be noted that horse remains were absent from subsequent strata (Hellwing, Feig, 1989: 236–242). The faunal assemblage of Tel Michal illustrates that by the seventeenth century BC the domesticated horse can safely be identified in the Levantine faunal record. Tel Michal is important, as its emergence is contemporary with the advent of Hyksos rule in Lower Egypt and the introduction of the domesticated horse there.
Discussion
Horses are relatively obscure in the archaeological record of sites both before and after the Late Bronze Age advent of chariot warfare. At the biblical city of Shiloh, three E. caballus skeletal remains can be accounted for in Late Bronze Age levels but did not appear in later Hellenistic and Roman strata when their existence is well attested in the region (Hellwing, Sadeh, Kishon, 1993: 311, 314). Megiddo IV contains seventeen pillared buildings dating to the eighth century BC theorized to have functioned as stables. However, there is a distinct lack of E. caballus remains at Megiddo IV that suggests that these structures may have served another function (Cantrell, 2006: 630–640).
Domestic status is impossible to determine unless bitwear studies or conjunctional archaeological materials are found in situ to indicate as such, although it can be assumed that by the seventeenth century BC Levantine E. caballus remains are likely of domestic stock. So why there is a distinct presence of E. caballus/ ferus throughout the Levant, Iran, and Anatolia during the Holocene prior to well-documented use of domesticated horses for chariot warfare and as beasts of burden? The answer may lie in that small pockets of feral populations may have continually inhabited these areas long after they were assumed extinct.
If domesticated horses were dispersed into the Levant from the Eurasian steppe at the end of the 4th millennium BC, it is evident that E. caballus did not supersede E. asinus in the role of favored pack animal. Horses had already spread from the Eurasian steppe via Transcaucasia into Anatolia, Iran, and Syria by the start of the 3rd millennium BC, nonetheless for unknown reasons there was not a further dispersal of E. caballus into the literate societies of Mesopotamia to the south, the Harrapan civilization to the southeast, and Egypt to the southwest until the late 3rd–early 2nd millennium BC (Postgate, 1992: 161; Bökönyi, 1997; Emery, 1960). This may be due to the inherent issues and expense in keeping horses or the general unavailability of good grazing land. Horses would have been an especially unattractive option when other cheaper and more easily managed draft animals were already being utilized in these areas.
E. caballus was not prolific in the Near East until the invention of the chariot permitted the use of horses for long-distance trade and warfare at the end of the 3rd to early 2nd millennium BC. The earliest dated chariots are found in Sintashta settlements within the Eurasian steppe region near the Ural Mountains. Horse-drawn chariots and harnessing equipment feature commonly in Sintashta burials within the Volga-Don forest-steppe region ca 2000 BC (Kuznetsov, 2006). If the chariot was a Sintashta invention, there may be some credence to the proposed connection between Indo-European language and the spread of horsemanship. The Sintashta culture has been tentatively linked to the Indo-Aryan branch of IndoEuropean languages due to their burial practices, ritual feasting on horses, and other practices alluded to within the Rig Veda. This has led some archaeologists to propose that Sintashta communities spoke an early form of IndoAryan language (Kuzmina, 2001; Anthony, 2007: 409). If this was true, Indo-Europeans may have been responsible for the spread of chariotry, but as the archaeozoological evidence suggests, not for the dissemination of the domesticated horse.
The scarcity of E. caballus remains in the Levant throughout the Holocene may be explained by specific horse-keeping practices, comparative to treatment of other domesticated animals that were used solely for primary or secondary products. According to Caroline Grigson (pers. comm., 2014), rarity of E. caballus remains is commonplace in later periods post-dating the Late Bronze Age, even from sites that literary evidence suggest should have kept them. Most faunal samples are of animals that are butchered and consumed onsite; the absence of horse carcasses may be due to the fact that they were not eaten and any remains may therefore be located offsite. The problem of E. caballus scarcity may have to do with an oversight in archaeological method; if horses were being “taken out to pasture”, perhaps archaeozoologists are not looking in the right place.
