Уровни гормонов гипофизарно-тиреоидной оси и их связь с овариальными гормонами во время овуляторного цикла у молодых кур-несушек (Gallus domesticus L.)
")
Автор: Лебедева И.Ю., Митяшова О.С., Алейникова О.В., Монтвила Е.К.
Журнал: Сельскохозяйственная биология @agrobiology
Рубрика: Вопросы репродукции
Статья в выпуске: 6 т.59, 2024 года.
Бесплатный доступ
Известно, что гипоталамо-гипофизарно-тиреоидная ось участвует в эндокринном контроле функции яичников у млекопитающих, однако ее роль в регуляции развития овариальных структурных компонентов у птиц до сих пор не ясна. Наличие в яичнике птиц тиреоидных рецепторов, дейодиназ и клеточных транспортеров для тиреоидных гормонов указывает на функционирование путей, опосредующих влияние гормонов щитовидной железы на овариальные фолликулы. Завершающие стадии фолликулогенеза у птиц тесно сопряжены с овуляторным циклом, в течение которого происходят изменения в экспрессии различных регуляторных факторов, приводящих к изменению содержания в крови половых стероидных гормонов и чувствительности к ним фолликулярных клеток. Нами впервые проведен сравнительный анализ уровней тиреоидных гормонов и тиреотропного гормона на разных стадиях овуляторного цикла и охарактеризована их ассоциация с содержанием овариальных стероидных гормонов в крови домашней курицы (Gallus domesticus L.). Целью настоящего исследования было изучение возможного участия гормонов гипофизарно-тиреоидной оси в физиологических процессах, контролирующих овуляцию у кур-несушек. Для исследований были отобраны 15 кур породы Hisex White в возрасте 27-28 нед, несущих непрерывно не менее 7 яиц. Кровь брали через 1,5; 6,5; 11, 16 и 21 ч после овуляции. В плазме крови методом иммуноферментного анализа определяли концентрацию общего и свободного тироксина (Т4), общего и свободного трийодтиронина (Т3), реверсивного Т3 (rT3), тиреотропного гормона (ТТГ), а также прогестерона, тестостерона и эстрадиола-17β. Анализ содержания половых стероидных гормонов в крови кур подтвердил его преовуляторное повышение через 21 ч после предшествующей овуляции. У исследованных птиц не наблюдалось существенного изменения уровней общего Т4 и общего Т3 в течение овуляторного цикла. В то же время через 16-21 ч после овуляции содержание rT3 в крови было в 1,2-1,3 раза ниже (p function show_abstract() { $('#abstract1').hide(); $('#abstract2').show(); $('#abstract_expand').hide(); }
Куры-несушки, овуляторный цикл, половые стероидные гормоны, гипофизарно-тиреоидная ось, трийодтиронин
Короткий адрес: https://sciup.org/142244129
IDR: 142244129 | УДК: 636.5:591.1 | DOI: 10.15389/agrobiology.2024.6.1169rus
The levels of pituitary–thyroid axis hormones and their relationship with ovarial hormones during the ovulatory cycle of young laying hens (Gallus domesticus L.)
The hypothalamic-pituitary-thyroid axis is known to participate in the endocrine control of the ovarian function in mammals, but its role in regulating the development of ovarian structural components in birds is still unclear. The presence of thyroid receptors, deiodinases, and cellular transporters for thyroid hormones in the avian ovary indicates the functioning of pathways mediating the effects of thyroid hormones on ovarian follicles. The final stages of folliculogenesis in birds are closely associated with the ovulatory cycle, during which changes in the expression of various regulatory factors occur, leading to changes in the blood content of sex steroid hormones and the sensitivity of follicular cells to them. We have performed for the first time a comparative analysis of the levels of thyroid hormones and thyroid-stimulating hormone at different stages of the ovulatory cycle and characterized their association with the content of ovarian steroid hormones in the blood of the domestic hen (Gallus domesticus L.). The aim of current study was to investigate the possible involvement of hormones of the pituitary-thyroid axis in the physiological processes that control ovulation in laying hens. For the research, 15 White Hisex hens aged 27-28 weeks, continuously laying at least 7 eggs, were selected. Blood sampling was carried out at 1.5, 6.5, 11, 16 and 21 h after ovulation. In the blood plasma, the concentrations of total and free thyroxine (T4), total and free triiodothyronine (T3), reverse T3 (rT3), thyroid-stimulating hormone (TSH), as well as progesterone, testosterone and estradiol-17β were determined using enzyme immunoassay. The concentration of total and free thyroxine (T4), total and free triiodothyronine (T3), reverse T3 (rT3), thyroid stimulating hormone (TSH), as well as progesterone, testosterone and estradiol-17β were determined in the blood plasma using enzyme immunoassay. Analysis of the blood content of sex steroid hormones in hens confirmed the presence of their preovulatory increase 21 h after the previous ovulation. In the birds studied, no significant changes in total T4 and total T3 levels occurred during the ovulatory cycle. However, 16-21 h after ovulation, the blood content of rT3 was 1.2-1.3 times lower (p function show_eabstract() { $('#eabstract1').hide(); $('#eabstract2').show(); $('#eabstract_expand').hide(); }
Текст научной статьи Уровни гормонов гипофизарно-тиреоидной оси и их связь с овариальными гормонами во время овуляторного цикла у молодых кур-несушек (Gallus domesticus L.)
Исследования последних лет убедительно продемонстрировали, что гормоны щитовидной железы не только входят в комплекс ключевых метаболических регуляторов (1), но и оказывают плейотропное влияние на многие органы позвоночных, включая гонады (2, 3).
Общая активность тиреоидной системы определяется как синтезом, так и метаболической активацией/инактивацией тиреоидных гормонов. Хотя тироксин (Т4) является основным гормоном, который секретирует щитовидная железа, он обычно рассматривается как предшественник трийодтиронина (Т3), или прогормон (4). Трийодтиронин обладает примерно в 4 раза большей биологической активностью, чем Т4, но его кон-
∗ Работа выполнена при финансовой поддержке Российского научного фонда (проект 22-16-00149).
центрация в крови и период полураспада в несколько раз меньше. Процесс дейодирования Т4 происходит в основном в нетиреоидных тканях и катализируется дейодиназами трех типов (5). Дейодиназа 1-го типа (DIO1), экспрессируемая главным образом в печени, отвечает за конверсию Т4 в Т3, содержащийся в кровеносной системе, тогда как дейодиназа 2-го типа (DIO2) контролирует локальную (внутриклеточную) генерацию Т3. Кроме того, DIO1 может участвовать в превращении Т4 в неактивный реверсивный Т3 (rT3). При этом основным ферментом, катализирующим преобразование Т4 в rT3, служит дейодиназа 3-го типа (DIO3), которая также инактивирует Т3, превращая его в дийодтиронин (Т2) (5, 6).
Функционирование тиреоидной системы в целом представляет собой комплекс сложных, тонко регулируемых процессов, реализация которых зависит от множества факторов. Так, периферический метаболизм тиреоидных гормонов служит дополнительным/альтернативным механизмом поддержания их локальной (тканеспецифической) концентрации, необходимой для оказания физиологического воздействия на клетки-мишени (7). Влияние гормонов щитовидной железы также определяется экспрессией клетками ядерных тиреоидных рецепторов α и β (TR α и TR β ), которые опосредуют их геномные эффекты, и мембранных интегриновых рецепторов (ITG V β 3), через которые активируются различные внутриклеточные сигнальные пути (4, 8, 9). Кроме того, для связывания с ядерны-ми рецепторами Т3 и Т4 должны проникать в клетки посредством диффузии или, в случае некоторых типов клеток, с помощью мембранных транспортеров, таких как монокарбоксилатные транспортеры MCT8 и MCT10 или органический анионный транспортер OATP1 (2, 4, 10). При этом способностью проникать в клетки и связываться со специфическими рецепторами обладают только свободные Т4 и Т3 (то есть не связанные с транспортными белками), доля которых составляет менее 0,1 % от всех тиреоидных гормонов, содержащихся в кровеносной системе (4).
Твердо установлено, что гипоталамо-гипофизарно-тиреоидная ось участвует в эндокринном контроле функции яичников у млекопитающих, а любые ее патологии обусловливают нарушения репродуктивной функции (4, 11-13). Причем тиреоидные гормоны могут не только оказывать непосредственное влияние на яичник, но и модулировать регуляторное действие репродуктивных гормонов, в первую очередь половых стероидных гормонов (14, 15). В то же время у птиц роль гипоталамо-гипо-физарно-тиреоидной оси в регуляции развития овариальных структурных компонентов и ее связь с другими эндокринными системами, контролирующими репродукцию, до сих пор не ясна. Присутствие в яичнике птиц тиреоидных рецепторов, дейодиназ и клеточных транспортеров для тиреоидных гормонов указывает на наличие путей , опосредующих действие гормонов щитовидной железы на овариальные фолликулы (16-18). В ряде исследований была выявлена ассоциация генов, контролирующих функционирование тиреоидной системы, с яйценоскостью индеек и уток (19, 20). Кроме того, у некоторых видов птиц, включая кур, in vivo и in vitro обнаружена взаимосвязь компонентов тиреоидной системы с репродуктивными гормонами (17, 18, 21). Также нами было показано модулирующее влияние Т3 на пролиферативную активность культивируемых клеток из преовуляторных фолликулов кур-несушек (22). Тем не менее нет никакой информации об участии гормонов щитовидной железы в регуляции in vivo роста и развития фолликулов в яичнике птиц.
Завершающие стадии фолликулогенеза у птиц тесно сопряжены с овуляторным циклом, который находится под контролем преовуляторной 1170
волны лютеинизирующего гормона и овариальных стероидных гормонов (23-25). При этом в течение овуляторного цикла происходят изменения в овариальной экспрессии различных регуляторных факторов, включая фолликулярные рецепторы для репродуктивных и метаболических гормонов, что влияет на чувствительность к ним фолликулярных клеток (26-28). Связь тиреоидных гормонов с овуляторным циклом птиц до сих пор исследована не была.
Нами впервые проведен сравнительный анализ уровней тиреоидных гормонов и тиреотропного гормона на разных стадиях овуляторного цикла и охарактеризована их ассоциация с содержанием овариальных стероидных гормонов в крови домашней курицы ( Gallus domesticus L.).
Целью настоящей работы было изучение возможного участия гормонов гипофизарно-тиреоидной оси в физиологических процессах, контролирующих овуляцию у кур-несушек.
Методика. Исследования проводили на молодых курах-несушках породы Hisex White, которые находились в отдельном хорошо вентилируемом помещении с центральным отоплением в условиях физиологического двора (ФИЦ ВИЖ им. академика Л.К. Эрнста). Домашнюю птицу содержали в отдельных клетках для индивидуального мониторинга яйценоскости, при 12-часовом освещении в сутки. Куры имели неограниченный доступ к воде, кормление осуществляли комбикормом Purina Special («Nestle Purina PetCare», США) в соответствии с рекомендациями производителя. Эксперименты с птицей проводили согласно принципам ветеринарной медицинской этики и Европейской конвенции по защите позвоночных животных, используемых для экспериментов или в иных научных целях (ETS ¹ 123, Страсбург, 1986). Протокол исследований на курах-несушках был одобрен комиссией по биоэтике ФГБНУ ФИЦ ВИЖ им. академика Л.К. Эрнста (протокол ¹ 3 от 24.05.2022).
Для исследований отобрали 15 кур в возрасте 27-28 нед, у которых периоды ежесуточной кладки яиц составляли не менее 7 сут (то есть несущих непрерывно не менее 7 яиц в одном цикле яйцекладки). Ежесуточный мониторинг времени яйцекладки проводили с помощью видеосистемы. Время овуляции рассчитывали, исходя из того, что она происходит через 30 мин после снесения яйца (29). На основании этих данных рассчитывали средний интервал между овуляциями и ожидаемое время следующей овуляции для каждой птицы.
Образцы крови кур брали через 1,5; 6,5; 11, 16 и 21 ч после предшествующей овуляции с таким расчетом, чтобы последнее время взятия попадало на период преовуляторного повышения уровней стероидных гормонов (29). Кровь брали из подкрыльцовой вены в пробирки с гепарином натрия. После получения плазмы образцы аликвотировали, замораживали и хранили при - 30 ° С для анализа половых стероидных гормонов и при - 70 ° С для анализа тиреоидных гормонов и ТТГ.
Концентрацию гормонов в плазме крови измеряли одновременно во всех собранных образцах методом иммуноферментного анализа (ИФА) при использовании планшетного спектрофотометра Униплан (ЗАО «Пикон», Россия). Анализы проводили с использованием наборов следующих производителей: «DRG Instruments GmbH», Германия (общий и свободный Т4, общий и свободный Т3), «Diagnostics Biochem Canada, Inc.», Канада (rT3), «Cloud-Clone Corp.», Китай (ТТГ) и ООО «Хема», Россия (эст-радиол-17β, прогестерон, тестостерон). Все образцы исследовали в двух аналитических повторностях, при этом коэффициент вариации не превышал 16 %.
Полученные данные обрабатывали методом однофакторного дисперсионного анализа с повторными измерениями (one-way repeated measures ANOVA) при помощи программы SigmaStat 4.0 («Systat Software, Inc.», США). При этом время после овуляции служило повторяющимся фактором. Данные были представлены как средние значения ( M) и их стандартные ошибки (±SEM). Оценку различий между сравниваемыми средними значениями проводили на основании критерия Тьюки (Tukey’s test). Для анализа корреляционных связей использовали коэффициент Спирмена (Spearman coefficient).
Результаты. Анализ содержания половых стероидных гормонов в крови кур-несушек подтвердил наличие его преовуляторного повышения через 21 ч после предшествующей овуляции (рис.). При этом концентрация прогестерона возрастала в 5,6 раза (p < 0,001), тогда как концентрация тестостерона и эстрадиола-17β — только в 1,9-2,1 раза (p < 0,001) по сравнению с базальным уровнем.
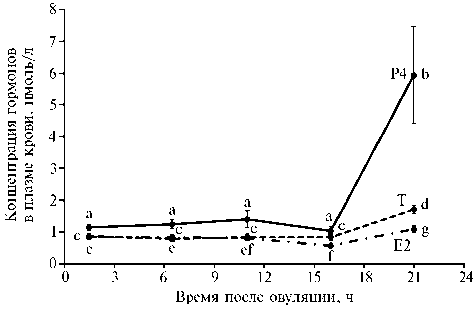
Концентрация прогестерона (P4) , тестостерона (T) и эстрадиола-17 β (E2) в крови кур ( Gallus domesticus L.) породы Hisex White в разные периоды овуляторного цикла ( n = 15, М ±SEM). Каждый образец исследовали в двух аналитических повторностях. Средние значения, помеченные индексами, не содержащими одинаковых букв, достоверно различаются (p < 0,0010,05).
Хотя концентрации общего Т4 и общего Т3 в крови исследованных птиц слегка варьировали в течение овуляторного цикла, уменьшаясь через 16 ч и увеличиваясь к 21-му ч, эти изменения не были существенными (табл. 1). В то же время через 16-21 ч после овуляции содержание rT3 в крови было в 1,2-1,3 раза ниже (p < 0,001-0,05), чем через 6,5-11 ч. Следовательно, снижение уровня неак- тивного rТ3 предшествовало преовуляторной волне половых стероидных гормонов.
1. Концентрация общего тироксина (Т4) , общего трийодтиронина (Т3) и реверсивного трийодтиронина (rТ3) в крови кур ( Gallus domesticus L.) породы
Hisex White в разные периоды овуляторного цикла ( n = 15, М ±SEM)
|
Время после овуляции, ч |
Концентрация гормонов в плазме крови, нмоль/л |
||
|
T4 |
T3 |
rT3 |
|
|
1,5 |
31,9±3,1 |
1,66 ± 0,25 |
0,324 ± 0,026 |
|
6,5 |
30,0±3,1 |
1,95 ± 0,32 |
0,348 ± 0,027a |
|
11 |
29,4±3,3 |
1,89 ± 0,30 |
0,361 ± 0,032b |
|
16 |
25,7±1,8 |
1,74 ± 0,21 |
0,305 ± 0,034c |
|
21 |
28,4±2,7 |
1,95 ± 0,20 |
0,277 ± 0,035d |
Примечание. Каждый образец исследовали в двух аналитических повторностях. Различия между группами статистически значимы: a,d p < 0,01; b,cp < 0,05; b,dp < 0,001.
Динамика изменения содержания ТТГ в крови кур была сходна с таковой для rT3 (табл. 2). Это содержание не изменялось между 1,5 и 11 ч после овуляции, а затем снижалось в 1,1 раза (p < 0,05) через 16 ч, оставаясь пониженным вплоть до преовуляторного пика овариальных гормонов.
Наблюдаемое снижение не могло быть следствием отрицательной обратной связи, существующей между ТТГ и свободным Т4 и свободным Т3 (4), поскольку уровни последних двух гормонов были относительно постоянными на протяжении всего овуляторного цикла.
-
2. Концентрация тиреотропного гормона (ТТГ) , свободного тироксина (fT4) и свободного трийодтиронина (fT3) в крови кур ( Gallus domesticus L.) породы Hisex White в разные периоды овуляторного цикла ( n = 15, Ì ±SEM)
-
3. Соотношение тиреоидных гормонов в крови кур ( Gallus domesticus L.) породы Hisex White в разные периоды овуляторного цикла ( n = 15, Ì ±SEM)
Время после овуляции, ч
Концентрация гормонов в плазме крови, нмоль/л
T4/T3 1
T3/rT3
fT4/fT3
-
4. Связь содержания тиреоидных и половых стероидных гормонов в крови кур ( Gallus domesticus L.) породы Hisex White в течение овуляторного цикла
|
Время после овуляции, ч |
Концентрация гормонов в плазме крови, пмоль/л |
||
|
ТТГ 1 |
fT4 |
fT3 |
|
|
1,5 |
14,0±1,2a |
8,07±1,20 |
2,29±0,19 |
|
6,5 |
14,3±1,0b |
7,16±1,06 |
2,56±0,25 |
|
11 |
14,1±1,2a |
8,14±1,19 |
2,45±0,26 |
|
16 |
12,8±1,3d |
7,05±0,93 |
2,50±0,21 |
|
21 |
12,2±1,4e |
7,68±1,21 |
2,26±0,23 |
Примечание. Каждый образец исследовали в двух аналитических повторностях. Различия между группами статистически значимы: a,ep < 0,05; b,dp < 0,05; b,ep < 0,01.
Во время преовуляторной волны половых гормонов (через 21 ч) также было выявлено повышение соотношения T3/rT3 в 1,5 раза (p < 0,05) по сравнению с таковым в период с 1,5 до 11 ч после овуляции (табл. 3). В то же время соотношение общих и свободных фракций Т4 и Т3 варьировалось несущественно.
Сравниваемые концентрации I Коэффициент корреляции r
|
Прогестерон |
Тестостерон |
0,693*** |
|
Тестостерон |
Эстрадиол-17 β |
0,531*** |
|
Прогестерон |
Эстрадиол-17 β |
0,377*** |
|
Общий трийодтиронин |
Прогестерон |
0,415*** |
|
Общий трийодтиронин |
Тестостерон |
0,503*** |
|
Тиреотропный гормон |
Общий трийодтиронин |
0,568*** |
|
Тиреотропный гормон |
Реверсивный трийодтиронин |
0,754*** |
|
Реверсивный трийодтиронин |
Общий трийодтиронин |
0,287* |
|
Тиреотропный гормон |
Свободный тироксин |
- 0,235* |
Примечание. Общее число проанализированных образов n = 15 (каждый образец исследовали в двух аналитических повторностях).
*, ** и *** Коэффициенты корреляции r статистически значимы соответственно при p < 0,05; p < 0,01 и p < 0,001.
Исследование показало умеренную позитивную взаимосвязь тестостерона с прогестероном и эстрадиолом-17β и слабую позитивную взаимосвязь (p < 0,001) между прогестероном и эстрадиолом-17β (табл. 4), что согласуется со сходной динамикой изменения уровней этих гормонов во время овуляторного цикла (см. рис.). При анализе связей между гормонами гипофизарно-тиреоидной оси и овариальными гормонами было выявлено наличие положительной корреляции (p < 0,001-0,01) между концентрацией в крови общего Т3 и содержанием прогестерона (слабой силы) и тестостерона (средней силы). Обнаружена умеренная положительная ассо- циация концентрации ТТГ и общего Т3 (r = 0,568; p < 0,001) и тесная ассоциация такой же направленности между уровнями ТТГ и реверсивного Т3 (r = 0,754; p < 0,001), тогда как концентрации T3 и rT3 были лишь слабо связаны друг с другом (r = 0,287; p < 0,05). При этом между содержанием ТТГ в крови и концентрацией свободного Т4 наблюдалась слабая отрицательная корреляция (p < 0,05).
Данные представленной работы показывают, что у кур-несушек преовуляторному пику половых стероидных гормонов предшествует уменьшение содержания в крови rT3. Хотя в печени кур присутствуют все три типа дейодиназ (30), отсутствие существенных изменений в уровне общего Т3 свидетельствует о снижении активности в печени DIO3, которая отвечает за конверсию Т4 в rT3, поступающий в кровеносную систему (5, 6). Наблюдаемому уменьшению концентрации rT3 в крови сопутствовало уменьшение концентрации ТТГ, что указывает на возможный вклад ТТГ в регуляцию активности DIO3. Действительно, у позвоночных печень — орган-мишень для ТТГ, тогда как у птиц последний рассматривается как модулятор активности дейодиназ 2-го и 3-го типа (31, 32).
Ранее K. Brady с соавт. (18) проводили сравнительное исследование содержания общего Т4 и общего Т3 в крови индеек во время преовулятор-ной волны репродуктивных гормонов и вне этой волны. Авторы показали (18), что у птиц со средней и высокой яйценоскостью уровни Т4 возрастают, а уровни Т3 понижаются во время преовуляторного пика гормонов. Однако этих данных недостаточно для определения направленности сдвига баланса тиреоидных гормонов в крови, поскольку не учитывался такой фактор, как rT3. Согласно общепринятому мнению, rT3 не обладает биологической активностью, однако он способен конкурировать с Т3 за связывание с мембранными тиреоидными рецепторами и, как следствие, ингибировать негеномные эффекты активного тиреоидного гормона (4, 6). Поэтому в клинической практике в последние годы предлагается рассматривать повышение концентрации rT3 и понижение соотношения T3/rT3 как признак латентного гипотиреоза (33).
В нашем текущем исследовании при использовании кур с высокой яйценоскостью и сравнении нескольких стадий овуляторного цикла не было обнаружено значимых изменений концентрации Т4 или Т3. В то же время анализ такого показателя, как уровень rT3, играющего ингибиторную роль в реализации влияния Т3 на клетки-мишени, выявил его снижение уже на стадии, предшествующей преовуляторной волне. Кроме того, постепенный рост соотношения T3/rT3, особенно выраженный через 21 ч после овуляции, указывает на сдвиг баланса в сторону наиболее активного тиреоидного гормона на завершающей стадии овуляторного цикла. Учитывая способность Т3 влиять на стероидогенную и пролиферативную активность клеток преовуляторных фолликулов кур (17, 22), можно предположить, что такой сдвиг будет приводить к усилению воздействия Т3 на эти фолликулы перед овуляцией.
В нашей работе, как и в других исследованиях, наблюдалось преову-ляторное повышение содержания в крови всех трех овариальных гормонов, однако у кур только прогестерон и тестостерон связаны с процессом овуляции (29, 34). Обнаруженная корреляция умеренной и слабой силы между уровнем общего Т3 и концентрацией прогестерона и тестостерона у кур указывает на определенную сопряженность их вариабельности во время овуляторного цикла, несмотря на отсутствие статистически значимых изменений уровня Т3. Этот вывод согласуется с многочисленными данными о наличии взаимосвязи между тиреоидной системой и репродуктив-1174
ными гормонами у позвоночных (14, 15).
Следует отметить отсутствие существенных изменений содержания в крови свободных фракций Т4 и Т3, а также их ассоциации с концентрацией половых стероидных гормонов. Тем не менее такие показатели могут не отражать локальный (тканевой) уровень этих гормонов, который зависит от активности дейодиназ в яичнике (6). Кроме того, свободный Т4, попадая в клетки-мишени посредством диффузии или использования мембранных транспортеров, превращается с помощью внутриклеточной DIO2 в Т3, который может оказывать эффекты через свои ядерные рецепторы, или с помощью внутриклеточной DIO3 — в rT3, обладающий низкой аффинностью к этим рецепторам (4). Следовательно, для характеристики степени воздействия тиреоидных гормонов на преовуляторные фолликулы кур необходимо исследовать экспрессию этих компонентов тиреоидной системы в фолликулярных клетках в динамике овуляторного цикла.
Сильная корреляционная связь, обнаруженная между содержанием в крови ТТГ и реверсивного Т3, свидетельствует в пользу нашего предположения о влиянии ТТГ на активность в печени DIO3, контролирующей конверсию Т4 в rT3. Кроме того, наличие умеренной ассоциации между уровнями ТТГ и общего Т3 согласуется с мнением ряда исследователей о роли ТТГ у птиц как модулятора активности DIO2, отвечающей за превращение Т4 в Т3 (30, 31). В то же время слабая отрицательная корреляция между концентрацией ТТГ и свободного Т4 указывает на лишь незначительный вклад отрицательной обратной связи между этими гормонами в вариабельность ТТГ.
Таким образом, у молодых кур-несушек снижение концентрации в крови тиреотропного гормона и реверсивного трийодтиронина предшествовало преовуляторной волне овариальных стероидных гормонов, тогда как во время этой волны происходил сдвиг баланса между общим и реверсивным трийодтиронином в сторону его активной формы. Обнаружено, что через 16-21 ч после овуляции содержание rT3 в крови птиц было в 1,21,3 раза ниже (p < 0,001-0,05), чем через 6,5-11 ч. Уровень ТТГ в крови не изменялся между 1,5 и 11 ч после овуляции, после чего происходило его снижение в 1,1-1,2 раза (p < 0,01-0,05). Во время преовуляторной волны половых гормонов (через 21 ч) выявлено повышение соотношения T3/rT3 в 1,5 раза (p < 0,05) по сравнению с таковым в период с 1,5 до 11 ч после овуляции. При этом во время овуляторного цикла наблюдалась положительная корреляция между концентрацией в крови общего Т3 и содержанием прогестерона ( r = 0,415; p < 0,001) и тестостерона ( r = 0,503; p < 0,001), что указывает на взаимосвязь (от слабой до умеренной) тиреоидной и репродуктивной систем. В целом полученные данные указывают на то, что во время преовуляторного пика половых стероидных гормонов у кур-несушек происходит изменение баланса гормонов гипофизарно-тиреоидной оси, и это может приводить к модуляции их воздействия на яичник непосредственно перед овуляцией. Для более детального понимания участия тиреоидной системы в регуляции процессов, контролирующих овуляцию у кур-несушек, необходимы дальнейшие исследования, чтобы охарактеризовать изменения других компонентов этой системы (экспрессии ядерных и мембранных рецепторов, дейодиназ и мембранных транспортеров) в процессе овуляторного цикла.
