Влияние цианобактерии Nostoc paludosum и ее экзометаболитов на рост ячменя

Автор: Фокина Анна Ивановна, Домрачева Людмила Ивановна, Огородникова Светлана Юрьевна
Журнал: Принципы экологии @ecopri
Рубрика: Оригинальные исследования
Статья в выпуске: 3 (33) т.8, 2019 года.
Бесплатный доступ
Изучен химический состав и влияние экзометаболитов почвенной цианобактерии Nostoc paludosum 18 на растения ячменя сорта Изумруд. Экзометаболиты из культуральной жидкости экстрагировали гексаном и четыреххлористым углеродом. Определение состава органических веществ в изучаемых субстратах проводили методом высокоэффективной жидкостной хроматографии на хроматографе Shimadzu LC-20, серия Prominence с диодноматричным детектором и газовой хроматографии на хроматографе GC-2014 Shimadzu с детектором TCD-2014 (Япония). Интенсивность перекисного окисления липидов в клетках ячменя оценивали по накоплению малонового диальдегида с использованием спектрофотометра SPEKOL 1300 (Analytik Jena, Германия). Цианобактериальная суспензия, культуральная жидкость и вытяжки из нее были близки по составу и количеству органических веществ и содержали такие соединения, как фитоин, фитофлуин, N-ацетилглюкозамин, пептидогликан муреин, ликопин, лютеин, каурен (предшественник гиббереллина), витамин А, провитамины, цианопептид борофицин. Стимулирующее действие экзометаболитов N. paludosum на линейный рост надземных органов и корней ячменя в основном проявляется в первые трое суток его роста. Цианобактериальные экзометаболиты снижают интенсивность перекисного окисления липидов в клетках растения, что свидетельствует об их антиоксидантной активности. Экзометаболиты цианобактериальной суспензии и гексановой вытяжки оказывают протекторное действие на растения, нивелируя ростингибирующий эффект ионов меди(II) на высоту проростков. Концентрация ионов меди(II) составляла 3 мг/кг субстрата для выращивания пшеницы. Выращивание ячменя на цианобактериальной суспензии и вытяжке экзометаболитов четыреххлористным углеродом приводит к накоплению цианопептида борофицина в тканях ячменя в количестве 0.0008-0.001 мкг/г сухой массы.
Цианобактерии, экзометаболиты, борофицин, ячмень
Короткий адрес: https://sciup.org/147231257
IDR: 147231257 | УДК: 579.69;
Influence of cyanobacterium Nostoc paludosum and its exometabolites on barley growth
The chemical composition and influence of exometabolites of the soil cyanobacterium Nostoc paludosum 18 on the barley plants of Izumrud variety were studied. Exometabolites from the culture liquid were extracted with hexane and carbon tetrachloride. Organic substances in the studied substrates were determined by high-performance liquid chromatography on the Shimadzu LC-20 chromatograph, Prominence series with a diode-matrix detector and gas chromatography on the GС-2014 Shimadzu chromatograph with a TCD-2014 detector (Japan). The intensity of lipid peroxidation in barley cells was studied by the accumulation of malondialdehyde using SPEKOL 1300 spectrophotometer (Analytik Jena, Germany). Cyanobacterial suspension, culture liquid and extracts from it were similar in composition and quantity of organic substances and contained such compounds as phytoene, phytofluin, N-acetylglucosamine, peptidoglycan murein, lycopene, lutein, kauren (a precursor of gibberellin), vitamin A, provitamins, peptide borofitsin. The stimulating effect of N. paludosum exometabolites on linear growth of barley aboveground organs and roots mainly manifests itself in the first three days of their growth. Cyanobacterial exometabolites reduce the intensity of lipid peroxidation in plant cells, which indicates their antioxidant activity. Exometabolites of cyanobacterial suspension and hexane extract have a protective effect on plants, leveling the growth- inhibitory effect of copper (II) ions on the height of seedlings. The concentration of copper (II) ions was 3 mg / kg of the substrate for growing wheat. Cultivation of barley on cyanobacterial suspension and extraction of exometabolites with carbon tetrachloride results in the accumulation of peptide borofitsin in barley tissues in the amount of 0,0008-0,001 µg / g of dry weight.
Текст научной статьи Влияние цианобактерии Nostoc paludosum и ее экзометаболитов на рост ячменя
К числу свойств цианобактерий (ЦБ), которые можно использовать в биотехнологическом аспекте, относится их способность синтезировать биологически активные вещества, выделяемые в окружающую среду в виде экзометаболитов. ЦБ выделяют свои метаболиты не только после отмирания клеток, но и в процессе нормальной физиологической деятельности. Выделение метаболитов ЦБ является для них одним из приспособлений к изменениям внешних условий (Сакевич, 1985). Первые детальные обзоры о химической природе подобных веществ в русскоязычной литературе обобщены в монографиях (Сиренко, Козицкая, 1988; Ан-дреюк и др., 1990). Однако в данных обзорах, как и в более поздних исследованиях, в основном приводятся сведения о структуре и свойствах, в том числе токсиногенной агрессивности, веществ, образуемых преимущественно водными формами ЦБ (Румянцев, Крюков, 2012; Белых и др., 2015; Voloshko et al., 2008; Wejnerowski et al., 2018; Rott et al., 2018). Данных же об экзогенных метаболитах почвенных ЦБ не так много. При этом почвенные ЦБ в течение многих лет используются в качестве агентов становления супрес-сивности химически и биологически загрязненных почв, а также для инокуляции семян различных сельскохозяйственных, лесных и декоративных культур (Домрачева, 2005; Домрачева и др., 2009). Полученные результаты о положительном влиянии интродукции ЦБ на повышение урожайности растений, подавлении активности фитопатогенов, их ремедиационной активности в почвах, загрязненных ионами тяжелых металлов, пестицидами, нефтепродуктами, требуют дальнейшего глубокого изучения химического состава экзометаболитов ЦБ. В частности, высокая метаболитическая активность и экологический потенциал отмечен у ЦБ рода Nostoc (Огородникова и др., 2010; Фокина и др., 2015; Řezanka, Dembitsky, 2006).
Цель данной работы – выявить состав и влияние экзометаболитов почвенной цианобактерии Nostoc paludosum Kütz 18 на рост и развитие растений ячменя сорта Изумруд в присутствии сульфата меди(II) и без него.
Материалы
В работе использовали:
– двухмесячную культуру ЦБ N. paludosum 18 с титром 3^107 кл./см3, выращенную на жидкой среде Громова № 6 без азота;- куль- туральную жидкость (КЖ) N. paludosum 18; – ячмень яровой сорта Изумруд. Выведен при обработке семян ячменя сорта Биос-1 биологическим препаратом «Агат 25 К».
Для эксперимента культуру ЦБ гомогенизировали в течение 2 минут с помощью гомогенизатора марки HG-15А-Set-A (DAIHAN Scientific, Ю. Корея) при 30 тыс. об./мин. Далее использовали суспензию ЦБ, ее КЖ (отделяли от микроорганизмов центрифугированием) и вытяжки из КЖ, приготовленные экстрагированием веществ из КЖ в гексан (С6Н) и четыреххлористый углерод (ЧХУ, ССl4). Экстракция гексаном – один из самых распространенных способов выделения биологически активных веществ (каротина, каротиноидов, токоферолов, флавоноидов и т. д.) из образцов растительного происхождения химически индифферентным нейтральным органическим растворителем (Ивкова, Петрова, 2012). Четыреххлористый углерод – зарекомендовавший себя неселективный экстрагент, способный растворять многие не только мономеры, но и соединения - полимеры (Химическая энциклопедия..., 2012). Экстракция проведена в соотношении КЖ : экстрагент, равном 1 : 1, порциями с последующим их соединением.
Определение состава органических веществ в КЖ, вытяжках из нее и растительных тканях. Определение состава органических веществ в изучаемых субстратах проводили методом высокоэффективной жидкостной хроматографии на хроматографе Shimadzu LC-20, серия Prominence с диодноматричным детектором и газовой хроматографии на хроматографе GC-2014 Shimadzu с детектором TCD-2014 (Япония).
Методы
Исследование влияния экзометаболитов почвенной цианобактерии N. paludosum на рост и развитие проростков ячменя
Фильтровальную бумагу пропитывали исследуемыми субстанциями (суспензия ЦБ, КЖ, вытяжки С6Н14 и CC14 из КЖ), высушивали в термостате до постоянной массы при 37 °С, помещали в стерильные чашки Петри и увлажняли дистиллированной водой таким образом, чтобы фильтр был увлажнен настолько, что последующая капля воды не впитывается в бумагу. Таким образом поддерживали влажность фильтра на максимально возможном уровне, ежедневно добавляя необходимое до полной влагоем-кости количество воды с помощью пипетки Пастера. На увлажненные фильтры раскла- дывали по 20 зерновок в каждую чашку Петри. Опыт проведен в трехкратной повторности для каждого варианта. На третьи и пятые сутки определяли высоту проростков и длину самого длинного корня. Для оценки влияния субстанций на развитие проростков ячменя сорта Изумруд использовали индекс роста (I) (Андреева, Кожевин, 2014):
I = (R + P ) · D, где I – индекс роста;
R + P – суммарные значения длин корней и проростков соответственно, см;
D – доля проросших семян, %.
Исследование влияния экзометаболитов почвенной цианобактерии N. paludosum на рост и развитие 7-суточных растений ячменя без зерновок
Фильтровальную бумагу помещали в чашки Петри и увлажняли стерильной дистиллированной водой до полной влагоемкости фильтра. Влажность фильтра поддерживали ежедневным внесением воды на фильтр пипеткой Пастера. На увлажненные фильтры раскладывали заранее обработанные в течение пяти минут 75 %-ным этиловым спиртом зерновки ячменя. На 7-е сутки проращивания отделяли зерновки от проростков пинцетом, чтобы исключить питание растений веществами, содержащимися в эндосперме зерновок.
Проростки без зерновок помещали в чашки Петри на фильтровальную бумагу (предварительно пропитанную раствором СuSO4; суспензией ЦБ; суспензией ЦБ + CuSO4; КЖ; КЖ + CUSO4; C6HM; C6HM + CUSO4; CCI4; CCI4 + CuSO4, высушенную до постоянной массы и перед раскладкой растений увлажненную стерильной водой). В контрольном варианте фильтры смачивали дистиллированной водой. В вариантах с сульфатом меди(II) была создана концентрация Си2+ 3 мг/кг субстрата, что соответствует предельно допустимой концентрации подвижных форм меди в почве. В каждом варианте было по 30 растений (n = 3). На 3-и сутки экспозиции проростков без зерновок определяли высоту проростков и длину самого длинного корня. Интенсивность перекисного окисления липидов (ПОЛ) в ячмене оценивали по накоплению малонового диальдегида (МДА), который образуется в растительных тканях в процессе ПОЛ при реакции с тиобарбитуровой кислотой (Лукаткин, 2002). Содержание МДА оценивали фотометрически с использованием спектрофотометра SPEKOL 1300 (Analytik Jena, Германия). Определяли содержание борофицина (Merken, Beecher, 2000) и меди в проростках (Сборник методик..., 2004). На протяжении всего эксперимента чашки с растениями находились в климатостате с температурой в ночное время +12–13 °С, в дневное – +21–23 °С.
Статистическая обработка и построение графиков выполнено в программе Excel 2002 for Windows. На рисунках приведены средние арифметические величины и стандартная ошибка. Достоверность различий между двумя средними оценивали с использованием t-критерия Стьюдента.
Результаты
Экзометаболитный состав культуральной жидкости цианобактерии N. paludosum. В процессе жизнедеятельности ЦБ образуется ряд биологически активных веществ, которые выделяются в окружающую среду. В условиях водной культуры экзометаболиты ЦБ накапливаются в КЖ. Установлено, что вытяжки из КЖ и, соответственно, сама КЖ содержат такие вещества, как фитоин (0.5–0.8 мкг/дм3), фитофлуин (0.6-О.8 мкг/дм3), N-ацетилглюкозамин (порядка 2.5 мкг/дм3), пептидогликан муреин, антиоксиданты: ликопин (190–195 мкг/ дм3 – органические вытяжки и КЖ, 240–250 мкг/дм3 – в суспензии ЦБ) и лютеин (20–25 мкг/дм3). Обнаружены гормоны, предшественники гиббереллинов (каурен), витамин А и провитамины. Следует отметить, что в основном составы цианобактериальной суспензии, КЖ и вытяжки из нее по количеству определенных органических веществ существенно не отличаются. Единственной отличительной особенностью является высокое содержание ликопина в суспензии ЦБ. Теоретически большинство из этих веществ могут оказывать положительное влияние на развитие растений. Ранее было установлено, что N. muscorum вырабатывает цианопептид борофицин, обладающий антимикробной активностью (Banker, Carmeli, 1998; Swain et al., 2017). Вероятно, именно он может определять антифунгальную активность исследуемых субстанций. В нашем эксперименте концентрация данного цианопептида при его экстракции в соотношении субстанция:экстрагент, равном 1:1, во всех типах субстанций находилась в пределах 0.05–0.06 мкг/дм3. Наличие борофицина у ЦБ рода Nostoc, в частности у Nostoc linkia и N. spongiaeforme, было отмечено в 1994 г. (Hemscheidt et al., 1994). Данный цианопептид биологически активен, поэтому необхо- димо исследовать возможность его миграции в растения.
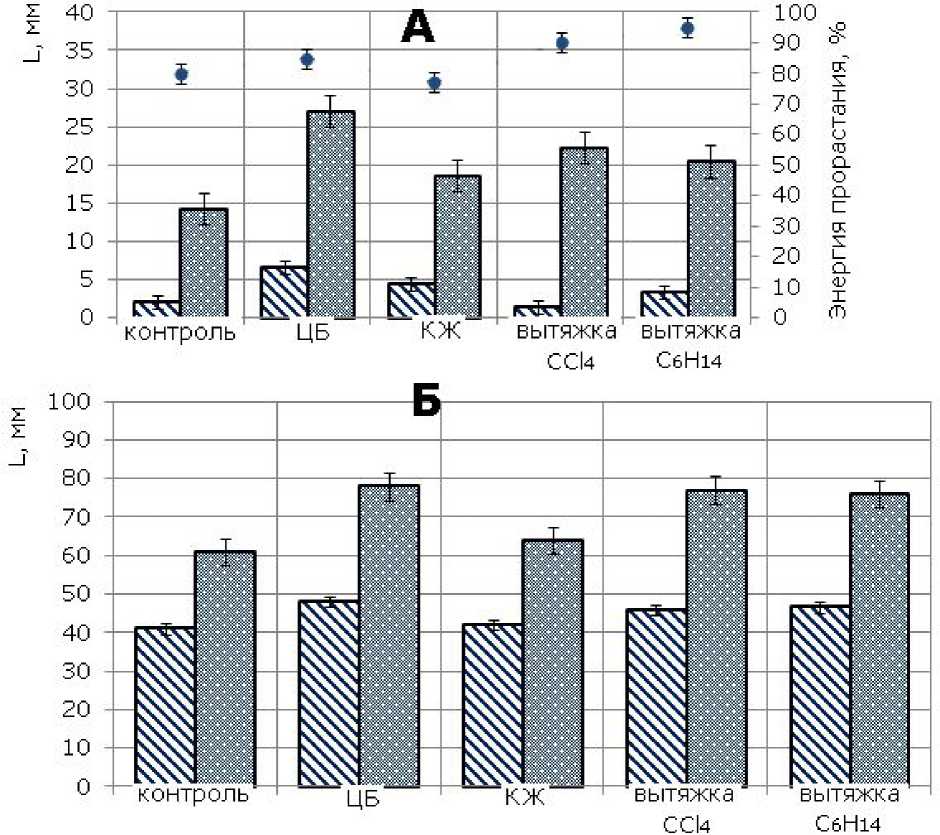
Влияние экзометаболитов цианобактерии N. paludosum на развитие проростков ячменя. Экзометаболиты ЦБ являются биологически активными соединениями, которые оказывают влияние на процессы прорастания семян, роста и развития проростков. На третьи сутки экспозиции ячменя на фильтрах, обработанных исследуемыми субстанциями, наибольшую энергию прорастания семян наблюдали в вариантах, выращиваемых на вытяжках С6Н14 (95 %), СС14 (90 %) и ЦБ (85 %) (рис. 1А). Достоверный ризогенный эффект, по сравнению с контролем, установлен во всех вариантах, проростков – в вариантах с ЦБ и КЖ. Максимальное стимулирующее действие на рост ячменя по обоим показателям оказала суспензия ЦБ. В этом варианте отличие от контроля составило почти 100 % как по длине корней, так и по высоте проростков.
К пятым суткам нивелируется стимулирующий эффект КЖ, хотя в остальных вариантах по-прежнему показатели высоты проростка и длины корня превышают данные показатели в контроле на 10–15 % и 33–37 % соответственно (рис. 1Б).
Вариант
ЕИПроростки ^Корни
♦-Энергия прорастания, %
Рис. 1. Влияние цианобактрии N. paludosum и ее экзометаболитов на морфометрические параметры ячменя сорта Изумруд (А – 3-е сутки, Б – 5-е сутки). «*» – результаты достоверно отличаются от контроля при Р > 0.95
Fig.1. The influence of cyanobacterium N.Paludosum and its exometabolites on morphometric parameters of barley of Izumrud variety (A - the 3rd day, B - the 5th day), "*" - results are reliably different from the controls at P >0.95
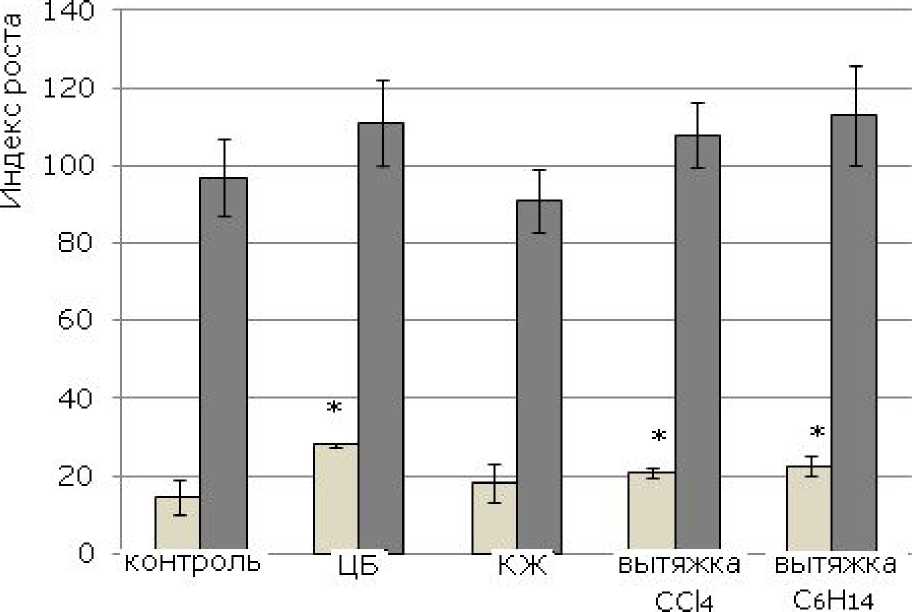
Использование показателя индекса роста (рис. 2) для иллюстрации наблюдаемых эффектов позволило выявить интегральную характеристику влияния исследуемых субстанций на рост и развитие растений ячменя. Использование цианобактериальной суспен зии, вытяжек С6Н14 и СС14 в целом оказывает ростстимулирующий эффект, что выявлено на 3-суточных проростках. К пятым суткам
опыта данная тенденция сохраняется, однако достоверно судить о стимуляции роста уже не удается.
□ 3-е сутки ОБ-е сутки Вариант
Рис. 2. Влияние цианобактерии N. paludosum и ее экзометаболитов на индекс роста растений ячменя сорта Изумруд. «*» – результаты достоверно отличаются от контроля при Р > 0.95 ( n = 60)
Fig. 2. The effect of cyanobacterium N. paludosum and its exometabolites on the plant growth index of barley Izumrud variety. "*" - results are reliably different from controls at P >95 (n=60)
Таким образом, установлено, что комплекс веществ всех субстанций стимулирует рост корней на ранних стадиях развития ячменя. При этом цианобактериальная суспензия обладает наибольшим ростстимулирую-щим действием.
Оценка влияния исследуемых веществ на рост и развитие растений ячменя, освобожденных от зерновок. Действие цианобактериальных метаболитов на рост ячменя изучали в опыте с 7-суточными проростками судаленными зерновками. 7-суточные растения без зерновок переносили в чашки Петри на фильтровальную бумагу, предварительно обработанную изучаемыми субстанциями. После 3-суточной экспозиции определяли морфометрические показатели, содержание борофицина в проростках, меди - в проростках и корнях.
Высота проростков. Наибольший рост-стимулирующий эффект по отношению к высоте проростков установлен в вариантах с вытяжкой C 6 H 14 (95 ± 3 мм) и суспензией ЦБ (94 ± 5 мм) против 90 ± 4 мм в контроле. Однако достоверного отличия между величинами данного параметра у всех вариантов без добавления сульфата меди(11) не обнаружено (рис. 3).
Длина корней. Выращивание 7-суточных растений ячменя без зерновок на средах, содержащих ЦБ и их метаболиты, привело к торможению роста корневой системы (см. рис. 3). Данное явление можно рассматривать как реакцию на стресс, направленную на адаптацию растений к условиям среды выращивания, или отсутствие необходимости к увеличению линейных размеров корней, обеспечивающему полноценное пита- ние растения. В варианте, где метаболиты ное отставание роста корней по сравнению с отсутствовали, ионы меди(II) в количестве, контролем равном 3 мг/кг субстрата, вызвали достовер-
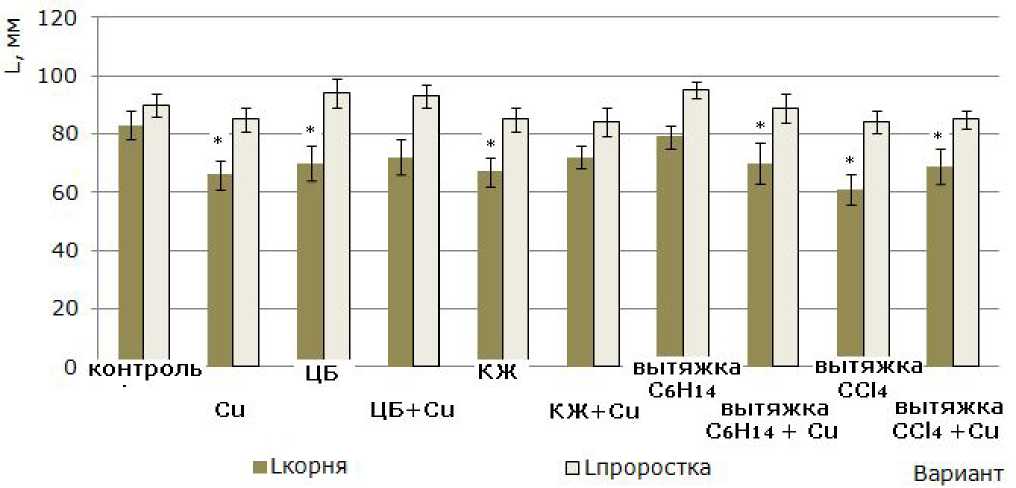
Рис. 3. Влияние N. paludosum, ее экзометаболитов и меди на рост проростков ячменя после удаления зерновок. «*» – результаты достоверно отличаются от контроля при Р > 0.95 ( n = 30)
Fig. 3. The effect of N. paludosum , its exometabolites and copper on the growth of barley seedlings after the removal of grains. "*" - results are reliably different from controls at P >0.95 (n=30)
Влияние компонентов культуральной жидкости цианобактерии на накопление меди корнями и проростками ячменя. Обнаружение меди в растениях вариантов без добавления соли металла является следствием того, что Cu – микроэлемент, входящий в состав зерновок в естественном состоянии. Среднее содержание меди в растениях, по Виноградову, составляет 2 мкг/г (Ягодин и др., 2002), допустимое остаточное количество – 10 мкг/г (Найштейн и др., 1987). В данные параметры вполне вписывается значение содержания меди не только контрольного варианта, но и вариантов с КЖ, вытяжками C 6 H 14 , СС1 4 (рис. 4). В остальных вариантах при благополучии с содержанием меди в проростках несколько повышено количество меди в корнях. При этом добавление сульфата меди(11) приводит к тенденции увеличения накопления металла.
Влияние компонентов культуральной жидкости на накопление борофицина в проростках ячменя. С одной стороны, цианопептид борофицин может вызывать отравления у млекопитающих, играя, таким образом, отрицательную роль, с другой – обладает антимикробными свойствами по отношению к фитопатогенам. Интерес, проявляемый к пептидам в последние годы, связан как с возможностью использования генов микроорганизмов, отвечающих за выработку вещества, для создания устойчивых форм сельскохозяйственных растений, так и с перспективностью их применения для разработки лекарственных препаратов нового поколения (Кокшарова, 2010; Одинцова и др., 2012).
Установлено, что борофицин накапливается в проростках при выращивании ячменя на суспензии ЦБ и вытяжке Сс1 4 из КЖ в количестве, равном 0.0008–0.001 мкг/г сухой массы. В проростках остальных вариантов его не обнаружено. При этом борофицин присутствует практически в одинаковом количестве (0.04 мкг/дм3) во всех субстанциях. Логично, что такая особенность связана с комплексом факторов как со стороны растения, так и силой действия комплекса веществ субстанций, на которых растения выращивали. Однозначного ответа пока нет.
Влияние экзометаболитов на перекисное окисление липидов в листьях ячменя. Перекисное окисление липидов - окислительная деградация липидов в растениях происходит в основном под действием свободных радикалов и является одним из симптомов окислительного стресса в растительных клетках (Apel, Hirt, 2004). В норме ПОЛ поддерживается на определенном уровне за счет эффективной работы систем анти-
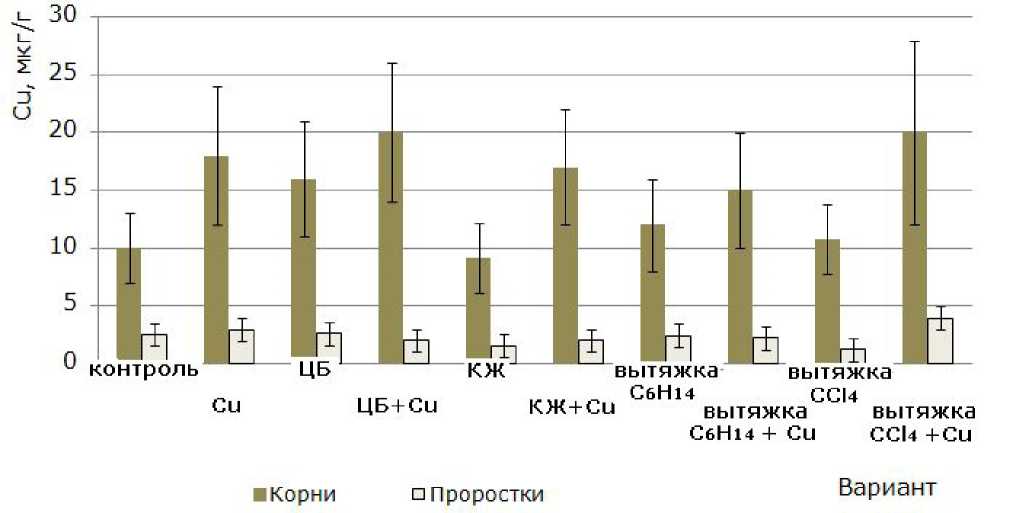
Рис. 4. Содержание меди в проростках и корнях 10-суточных растений ячменя сорта Изумруд, освобожденных от зерновок ( n = 30; P = 0.95)
Fig. 4. The content of copper in the seedlings and roots of 10-day-old barley of Izumrud variety, after the removal of grains (n=30; P =0.95)
оксидантной защиты (антиокислительные ферменты, вещества с антиоксидантными свойствами) (Blokhina et al., 2003). В стрессовых условиях изменение интенсивности процессов ПОЛ может свидетельствовать как о патологических состояниях (Лелевич, 2009), так и о процессах адаптации.
Выращивание проростков ячменя на испытуемых субстратах приводило к достоверному, по сравнению с контролем (вода), снижению накопления продукта ПОЛ – малонового диальдегида (МДА) в клетках листьев (рис. 5). Низкая интенсивность процессов ПОЛ отмечалась в варианте с действием солей меди, что, возможно, связано с ходом адаптационных перестроек в клетках. Известно, что сразу после действия стресс-фактора в клетках происходит значительная активация антиоксидантных ферментов и накопление веществ с антиоксидантными свойствами. Все это приводит к снижению уровня активных форм кислорода, что сказывается на скорости протекания процессов ПОЛ в клетках. Известно, что фосфорорганический ксенобиотик – метилфосфоновая кислота и ЦБ – N. muscorum вызывают снижение уровня МДА в клетках листьев ячменя (Коваль, Огородникова, 2014), низкие температуры, также могут приводить к уменьшению уровня МДА в клетках листьев табака
(Попов и др., 2010). Снижение интенсивности ПОЛ в клетках ячменя, выращенных на испытуемых субстратах, возможно, обусловлено действием на них экзометаболитов ЦБ, в составе которых присутствуют вещества с выраженными антиоксидантными свойствами – ликопин, лютеин, витамин А и др.
Заключение
В культуральной жидкости цианобактерии N. paludosum методом ВЭЖХ обнаружены вещества, которые могут обладать биологической активностью (ликопин, лютеин – антиоксиданты, витамин А и провитамины, предшественники гиббереллинов), способствующей росту ячменя. Также в составе культуральной жидкости обнаружен цианопептид борофицин. Цианобактерия N. paludosum 18 и еe экзометаболиты в большей степени влияют на развитие ячменя сорта Изумруд в первые трое суток его роста, оказывая стимулирующее действие, на линейный рост надземных органов и корней. Исследование показало, что присутствие цианобактерии N. paludosum и ее экзометаболитов приводит не только к ростстиму-лирующему эффекту, но и снижению интенсивности процессов перекисного окисления липидов в растительных клетках.
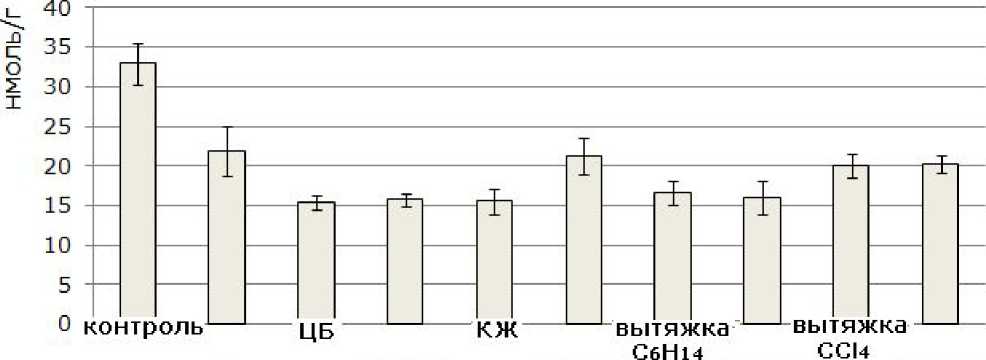
Си ЦБ+Си КЖ+Си вытяжка вытяжка
СбН14+Си CCh+Cu
Вариант
Рис. 5. Содержание малонового диальдегида в клетках листьев 10-суточных проростков ячменя сорта Изумруд, освобожденных от зерновок (пересчет на сырую биомассу)
Fig.5. The content of malonic dialdehyde in the leaf cells of 10-days-old seedlings of barley of Izumrud variety after the removal of grains (raw biomass equivalent)
Список литературы Влияние цианобактерии Nostoc paludosum и ее экзометаболитов на рост ячменя
- Андреева О. А., Кожевин П. А. Оптимизация естественного сообщества микроорганизмов почвы как способ создания микробных удобрений [Optimization of the natural community of soil microorganisms as a way to create microbial fertilizers] // Вестник Московского университета. Сер. 17: Почвоведение. 2014. № 4. С. 42-45.
- Андреюк Е. И., Коптева Ж. П., Занина В. А. Цианобактерии [Cyanobacteria]. Киев: Наукова думка, 1990. 200 с.
- Белых О. И., Гладких А. С., Сороковикова Е. Г., Тихонова И. В., Потапов С. А., Бутина Т. В. Сакситоксин-продуцирующие цианобактерии в озере Байкал [Saxitoxin-producing cyanobacteria in Lake Baikal] // Сибирский экологический журнал. 2015. № 2. С. 229-237.
- Домрачева Л. И. "Цветение" почвы и закономерности его развития ["Flowering" of the soil and the patterns of its development]. Сыктывкар, 2005. 336 с.
- Домрачева Л. И., Кондакова Л. В., Попов Л. Б., Зыкова Ю. Н. Биоремедиационные возможности почвенных цианобактерий (обзор) [Bioremediation capabilities of soil cyanobacteria (review)] // Теоретическая и прикладная экология. 2009. № 1. С. 8-17.
- Ивкова А. В., Петрова С. Н. Качественный анализ гексановых экстрактов листьев шиповника [Qualitative analysis of hexane extracts of wild rose leaves] // Известия вузов. Прикладная биохимия и биотехнология. 2012. № 2 (3). С. 158-159.
- Коваль Е. В., Огородникова С. Ю. Влияние цианобактерии Nostoc muscorum на устойчивость растений ячменя к действию метилфосфоновой кислоты [The effect of cyanobacterium Nostoc muscorum on the resistance of barley plants to the action of methylphosphonic acid] // Теоретическая и прикладная экология. 2014. № 2. С. 61-66.
- Кокшарова О. А. Применение методов молекулярной генетики и микробиологии в экологии и биотехнологии цианобактерий [Application of molecular genetics and microbiology in ecology and biotechnology of cyanobacteria] // Микробиология. 2010. Т. 79. № 6. С. 734-747.
- Лелевич В. В. Биологическая химия [Biological Chemistry]. Гродно: ГрГМУ, 2009. 275 с.
- Лукаткин А. С. Холодовое повреждение теплолюбивых растений и окислительный стресс [Cold damage to heat-loving plants and oxidative stress]. Саранск: Изд-во Мордов. ун-та, 2002. 208 с.
- Найштейн С. Я., Меренюк Г. В., Чегринец Г. Я. Гигиена окружающей среды и применение удобрений [Environmental health and fertilizer use]. Кишинев: Штиинца, 1987. 143 с.
- Огородникова С. Ю., Зыкова Ю. Н., Березин Г. И., Домрачева Л. И., Калинин А. А. Комплексная оценка состояния цианобактерии при воздействии различных поллютантов [Comprehensive assessment of cyanobacteria under the influence of various pollutants] // Теоретическая и прикладная экология. 2010. № 3. С. 47-51.
- Одинцова Т. И., Коростылева Т. В., Уткина Л. Л., Андреев Я. А., Славохотова А. А., Истомина Е. А., Пухальский В. А., Егоров Ц. А. Антимикробные пептиды пшеницы [Wheat antimicrobial peptides] // Вавиловский журнал генетики и селекции. 2012. Т. 16. № 1. С. 107-115.
- Попов В. Н., Антипина О. В., Трунова Т. И. Перекисное окисление липидов при низкотемпературной адаптации листьев и корней теплолюбивых растений табака [Lipid peroxidation during low-temperature adaptation of the leaves and roots of thermophilic tobacco plants] // Физиология растений. 2010. Т. 57. № 1. С. 153-156.
- Румянцев В. А., Крюков Л. Н. Особенности природы цианобактерий [Features of the nature of cyanobacteria] // Здоровье населения и среда обитания. 2012. № 2. С. 221-227.
- Сакевич А. И. Экзометаболиты пресноводных водорослей [Freshwater Algae Exometabolites]. Киев: Наукова думка, 1985. 199 с.
- Сборник методик измерений концентрации ионов меди, кадмия, цинка, висмута, марганца и никеля методом вольтамперометрии на вольтамперометрическом анализаторе "Экотест-ВА" [Collection of methods for measuring the concentration of copper, cadmium, zinc, bismuth, manganese and nickel ions using voltammetry using the Ecotest-VA volt-ampere analyzer]. М.: ООО "Эконикс-Эксперт", 2004. 61 с.
- Сиренко Л. А., Козицкая В. Н. Биологически активные вещества водорослей и качество воды [Biologically active substances algae and water quality]. Киев: Наукова думка, 1988. 256 с.
- Фокина А. И., Горностаева Е. А., Огородникова С. Ю., Зыкова Ю. Н., Домрачева Л. И., Кондакова Л. В. Адаптационные резервы почвенных природных биопленок с доминированием цианобактерий рода Phormidium [Adaptation potential of naturally occurring cynaobacterial biofilms dominated by phormidium ] // Сибирский экологический журнал. 2015. № 2. С. 842-851.
- DOI: 10.15372/SEJ20150604
- Химическая энциклопедия [Chemical encyclopedia] / Под ред. Н. С. Зефирова М.: Большая Российская энциклопедия, 1998. Т. 5. 783 с.
- Ягодин Б. А., Жуков Ю. П., Кобзаренко В. И. Агрохимия [Agrochemistry] / Под ред. Б. А. Ягодина. М.: Колос, 2002. 584 с.
- Apel K., Hirt H. Reactive Oxygen Species: Metabolism, Oxidative Stress, and Signal Transduction // Annu. Rev. Plant Biol. 2004. Vol. 55. P. 373-399.
- DOI: 10.1146/annurev.arplant.55.031903.141701
- Banker R., Carmeli S. Tenuecyclamides A-D, cyclic hexapeptides from the cyanobacterium Nostoc spongiaeforme var. tenue // Journal of Natural Products. 1998. Vol. 61. Issue 10. P. 1248-1251.
- DOI: 10.1021/np980138j
- Blokhina O., Virolainen E., Fagerstedt K. V. Antioxidants, Oxidative Damage and Oxygen Deprivation Stress: a Review // Annals of Botany. 2003. Vol. 91. Р. 179-194.
- DOI: 10.1093/aob/mcf118
- Hemscheidt T., Puglisi M. P., Larsen L. K., Patterson G. M. L., Moore R. E., Rios J. L., Clardy J. Structure and biosynthesis of borophycin, a new boeseken complex of boric acid from a marine strain of the blue-green alga Nostoc linckia // J. Org. Chem. 1994. Vol. 59. P. 3467-3471.
- Merken H. M., Beecher G. R. Liquid chromatographic method for the separation and quantification of prominent flavonoid aglycones // Journal of Chromatography A. 2000. Vol. 897. Issues 1-2. Р. 177-184.
- DOI: 10.1016/s0021-9673(00)00826-8
- Řezanka T., Dembitsky V. M. Metabolites produced by cyanobacteria belonging to several species of the family Nostocaceae // Folia Microbiologica. 2006. Vol. 51. Issue 3. P. 159-182.
- DOI: 10.1007/BF02932119
- Rott E., Pentecost A., Mareš J. Introduction: Recent developments in cyanobacterial research with special reference to aquatic habitats, molecular ecology and phylogenetic taxonomy // Hydrobiologia. 2018. Vol. 811. Issue 1. P. 1-6.
- DOI: 10.1007/s10750-017-3468-9
- Swain S. S., Paidesetty S. K., Padhy N. R. Antibacterial, antifungal and antimycobacterial compounds from cyanobacteria // Biomedicine & Pharmacotherapy. 2017. June. Vol. 90. P. 760-776. 10.1016 / j. biopha.2017.04.030
- DOI: 10.1016/j.biopha.2017.04.030
- Voloshko L., Safronova T., Pljusch A., Titova N., Kopecky J., Hrouzek P., Drabkova V. Toxins and other bioactive compounds produced by cyanobacteria in Lake Ladoga // Estonian Journal of Ecology. 2008. No 2. P. 100-110.
- DOI: 10.3176/eco.2008.2.02
- Wejnerowski Ł., Rzymski P., Kokociński M., Meriluoto J. The structure and toxicity of winter cyanobacterial bloom in a eutrophic lake of the temperate zone // Ecotoxicology. 2018. Vol. 27. Issue 6. P. 752-760. 10.1007 / s10646-018-1957-X.
- DOI: 10.1007/s10646-018-1957-X
