Влияние дегидратации и солевого рациона на динамику осморегуляторных гормонов у уток

Автор: Кучеров А.А., Пудовкин Н.А., Смутнев П.В.
Рубрика: Ветеринария
Статья в выпуске: 4 т.264, 2025 года.
Бесплатный доступ
В статье представлены результаты исследований по изучению влияния длительного обезвоживания и пищевого хлорида натрия на уровень аргинин-вазотоцина, пролактина, альдостерона и кортикостерона в крови у уток. Исследование проводили на птицах двух групп: контрольной (низкосолевой рацион + вода) и опытной (высокосолевой рацион + 0,5% раствор хлорида натрия). После девяти суток адаптации к рациону были установлены достоверные различия между группами по осмоляльности плазмы, концентрациям натрия и хлора, которые соответствовали уровню потребления хлорида натрия. Длительное обезвоживание вызвало прогрессирующее увеличение осмоляльности плазмы и концентраций натрия и хлора во всех группах, причем у птиц опытной группы эти показатели были достоверно выше. Концентрация калия оставалась относительно стабильной в группе с высоким потреблением натрия в период дегидратации. Исследование выявило значительное повышение уровня аргинин-вазотоцина, который демонстрировал прямую зависимость от уровня потребления хлорида натрия и сильную положительную корреляцию с осмоляльностью плазмы (r=0,958-0,987). Однако при обезвоживании секреция аргинин-вазотоцина достигала плато, несмотря на продолжающийся рост осмоляльности. Уровень пролактина достоверно повышался, демонстрируя тесную корреляцию с осмоляльностью плазмы (r=0,911-0,995) и концентрацией аргинин-вазотоцина (r=0,932-0,957). В отличие от этих гормонов, концентрация альдостерона значительно снижалась на фоне высокого потребления хлорида натрия и демонстрировала слабую отрицательную корреляцию с осмоляльностью и электролитами. Концентрация кортикостерона закономерно увеличивалась в ответ на дегидратацию. Полученные данные свидетельствуют о комплексных изменениях в осморегуляции и гормональном статусе уток при длительном обезвоживании и различном уровне потребления хлорида натрия.
Осморегуляция, дегидратация, аргинин-вазотоцин, утки, осмоляльность плазмы, пролактин, альдостерон, кортикостерон, электролиты (натрий, хлор, калий)
Короткий адрес: https://sciup.org/142246745
IDR: 142246745 | УДК: 619:616.36:577.1(470.46) | DOI: 10.31588/2413_4201_1883_4_264_57
Effect of dehydration and salt diet on dynamics of osmoregulatory hormones of ducks
This article presents results of a study examining the effects of prolonged dehydration and dietary sodium chloride on blood arginine vasotocin, prolactin, aldosterone, and corticosterone levels in ducks. The study was conducted on birds from two groups: a control group (low-salt diet + water) and an experimental group (highsalt diet +0.5% sodium chloride solution). After nine days of adaptation to the diet, significant differences were found between the groups in plasma osmolality, sodium and chloride concentrations, which corresponded to the level of sodium chloride intake. Long-term dehydration caused a progressive increase in plasma osmolality, sodium and chloride concentrations in all groups, with these values being significantly higher in the experimental group. Potassium concentration remained relatively stable in the high-sodium group during the dehydration period. The study revealed a significant increase in arginine vasotocin levels, which demonstrated a direct relationship with sodium chloride intake and a strong positive correlation with plasma osmolality (r=0.958-0.987). However, with dehydration, arginine vasotocin secretion reached a plateau, despite the continued increase in osmolality. Prolactin levels increased significantly, demonstrating a close correlation with plasma osmolality (r=0.911-0.995) and arginine vasotocin concentrations (r=0.932-0.957). In contrast to these hormones, aldosterone concentrations significantly decreased with high sodium chloride intake and demonstrated a weak negative correlation with osmolality and electrolytes. Corticosterone concentrations consistently increased in response to dehydration. These data indicate complex changes in osmoregulation and hormonal status in ducks during prolonged dehydration and varying levels of sodium chloride intake.
Текст научной статьи Влияние дегидратации и солевого рациона на динамику осморегуляторных гормонов у уток
Регуляция физиологических процессов в почках и дистальном отделе кишечника у птиц при обезвоживании и дефиците натрия хлорида осуществляется несколькими гормональными системами. Ключевую роль играют пептид аргинин-вазотоцин, пролактин и стероидные гормоны надпочечников - альдостерон и кортикостерон. Считается, что стероиды и пролактин участвуют в модуляции постренальной обработки мочи в кишечнике. Однако из -за недостатка сопоставимых данных in vivo, которые бы коррелировали плазменные концентрации этих гормонов с изменениями осмотического гомеостаза, их роль в осморегуляции птиц остается не до конца выясненной [3].
Целью исследования было определение динамики концентраций аргинин-вазотоцина, пролактина, альдостерона и кортикостерона в плазме крови в условиях продолжительного обезвоживания, сочетающегося с рационами, различающимися по содержанию хлорида натрия. Вариация уровня соли в рационе была предназначена для того, чтобы помочь разграничить влияние осмотических и волюмиче-ских (объемных) факторов.
Условия, материалы и методы. В исследовании использовали двенадцать уток пекинской породы. За неделю до начала эксперимента птиц перевели на диету с пониженным содержанием хлорида натрия и разместили индивидуально в клетках при 12-часовом световом режиме. Для минимизации стрессового воздействия были отобраны особи, которые ранее дважды успешно перенесли процедуру пункции сердца.
В первые сутки эксперимента у шести случайно выбранных уток отобрали исходные пробы крови. Затем всех птиц распределили на две опытные группы: контрольная группа (n=6) получала рацион с низким содержанием хлорида натрия и водопроводную воду, а опытная группа (n=6) – рацион с высоким содержанием хлорида натрия и 0,5% раствор хлорида натрия. Высокосолевой корм готовили путем добавления 1% хлорида натрия к базовому рациону, что повышало концентрацию натрия с 5 до 176 мкмоль/г при влажности корма около 12%.
Забор крови проводили на 3-е, 6-е и 9-е сутки адаптации к рационам. На 9-й сутки у птиц инициировали обезвоживание, удалив питьевую воду, и продолжили мониторинг с забором крови на 12-е, 15-е, 18-е, 21-е и 24-е сутки эксперимента. После 8-х суток в опытной группе пала одна птица, что сократило размер группы до пяти особей.
Кровь объемом 6-7 мл отбирали пункцией сердца в гепаринизированные пробирки в интервале между 08:00 и 08:30. Пробы немедленно охлаждали на льду и центрифугировали. Временной интервал от поимки птицы до завершения процедуры не превышал 3 минут (в среднем 75±5 секунд). Плазму сразу же аликвотировали: 1 мл для анализа аргинин-вазотоцина, остальные образцы хранили при 4 °C для последующего определения концентраций альдостерона, кортикостерона, электролитов и осмоляльности. Для анализа пробы пролактина (250 мкл) стабилизировали 0,1% азидом натрия.
Рассчитанные уровни потребления натрия составляли 0,2 и 15,3 ммоль/кг массы тела в сутки для рационов с низким (контроль) и высоким (опыт) содержанием хлорида натрия соответственно. Птицы группы с высокосолевым рационом и водопроводной водой потребляли примерно 7 ммоль натрия /кг в сутки.
Анализ данных проводили с использованием непараметрических методов: однофакторного дисперсионного анализа (тест Краскела-Уоллиса) и t-теста Манна-Уитни. Статистически значимыми считали различия при p<0,05.
Результаты и обсуждение. Исходные показатели (0 сутки) у шести случайно отобранных птиц, содержавшихся на низкосолевом рационе в течение 7 суток до эксперимента, составили: осмоляльность - 315± 4 мОсм, концентрация натрия - 142,01± 10,12 ммоль/л, хлора - 109,31±7,91 ммоль/л. Результаты исследований представлены в таблице 1.
Таблица 1 – Осмоляльность и концентрация электролитов плазмы крови уток
|
Показатель |
Сутки |
Осмоляльность (мОсм/л) |
Натрий (ммоль/л) |
Хлор (ммоль/л) |
Калий (ммоль/л) |
|
Контрольная группа |
|||||
|
Гидратация |
0 |
303,10±10,36 |
142,01±10,12 |
109,31±7,91 |
4,31±0,31 |
|
3 |
305,25±21,41 |
143,14±8,36 |
110,36±9,36 |
4,36±0,41 |
|
|
6 |
308,01±19,14 |
145,25±10,22 |
110,96±8,02 |
5,00±0,66 |
|
|
9 |
310,78±11,47 |
152,21±10,03 |
113,58±7,96 |
5,20±0,24* |
|
|
Дегидратация |
12 |
345,21±21,79* |
158,25±14,20 |
117,36±10,23 |
6,00±0,37* |
|
15 |
347,25±25,17* |
163,27±16,32 |
123,58±11,00 |
5,98±0,66* |
|
|
18 |
352,14±30,00* |
165,01±11,02 |
129,02±10,36* |
5,02±0,25* |
|
|
21 |
356,99±35,74* |
165,81±13,2* |
130,00±9,39* |
5,69±0,37* |
|
|
24 |
362,14±34,33* |
166,00±9,23* |
131,25±10,30* |
5,81±0,51* |
|
|
Опытная группа |
|||||
|
Гидратация |
0 |
305,12±25,01 |
151,01±6,21 |
113,95±10,36 |
5,29±3,01 |
|
3 |
329,23±31,01 |
152,01±7,95 |
115,98±9,35 |
5,36±0,31 |
|
|
6 |
350,21±21,99 |
155,31±9,31 |
117,98±7,36 |
5,37±0,66 |
|
|
9 |
354,36±31,00 |
159,01±10,03 |
119,35±10,91 |
5,31±0,12 |
|
|
Дегидратация |
12 |
360,25±19,97* |
163,25±12,81 |
126,35±11,91 |
5,40±0,41 |
|
15 |
361,21±25,87* |
171,20±15,00 |
139,25±12,03* |
5,34±0,39 |
|
|
18 |
362,21±21,91* |
173,01±10,91* |
140,99±10,36* |
5,21±0,54 |
|
|
21 |
370,21±34,21* |
177,31±13,36* |
141,02±8,07* |
5,63±0,21 |
|
|
24 |
378,25±31,21* |
178,00±12,92* |
144,36±9,96* |
5,61±0,10 |
|
Примечание: * - р<0,05 - достоверность различий относительно 0 суток
После девяти суток адаптации к рациону и перед обезвоживанием между двумя группами птиц наблюдали достоверные различия по осмоляльности плазмы, натрия и хлора. Эти различия соответствовали уровню потребления хлорида натрия.
Через три дня после удаления питьевой воды осмоляльность плазмы достоверно увеличилась на 13,8% у птиц в контрольной группе и на 18,1% у уток в группе с повышенным содержанием соли. Аналогичная динамика была отмечена для концентраций натрия и хлора. В период дегидратации не выявлено различий по осмоляльности у птиц контрольной группы, однако у птиц получавших высокосолевой рацион этот параметр был достоверно выше на 18,1-24,0% (р<0,05).
На протяжении последующих 12 суток дегидратации осмоляльность плазмы и концентрация натрия продолжали устойчиво возрастать в плазме крови птиц всех групп. К 24-м суткам значения у птиц каждой группе достоверно превышали показатели 10-х суток. Аналогичная динамика наблюдалась для концентрации хлоридов в плазме крови птиц контрольной (+1,0^ +20,1%) и опытной групп (1,8%...26,7%).
Исходный уровень калия у шести случайно отобранных птиц на низкосолевом рационе (0 сутки) составлял 4,31± 0,31 ммоль/л. К 9-м суткам эксперимента концентрация калия в плазме крови птиц контрольной группы возросла на 14%, достигнув 5,20±0,24 ммоль/л, что указывает на период стабилизации, длящийся не менее двух недель.
В условиях нормальной гидратации между группами птиц существовали значимые различия: к 8-м суткам концентрация калия у уток на высокосолевом рационе была на 10-13% ниже, чем у уток с повышенным содержанием соли. При этом внутри опытной группы птиц разница по концентрации калия в плазме крови была статистически незначимой.
В период дегидратации концентрация калия в плазме крови птиц оставалась стабильной в группе с повышенным потреблением натрия по сравнению с 8-м днем (табл. 1).
Наши результаты демонстрируют четкую ступенчатую зависимость: с увеличением уровня потребления хлорида натрия в рационе пропорционально росли осмоляльность плазмы и концентрации натрия и хлора. При этом концентрация калия была ниже у птиц на высокосолевом рационе, без существенных различий между группами.
Таблица 2 - Концентрации аргинин-вазотоцин, пролактина, альдостерона и кортикостерона в плазме
|
Показатель |
Сутки |
Аргинин-вазотоцин (нг/л) |
Пролактин (нг/л) |
Альдостерон (нг/л) |
Кортикостерон (нг/мл) |
|
Контрольная группа |
|||||
|
Гидратация |
0 |
8,17±0,13 |
28,39±1,97 |
165,00±9,24 |
1,10±0,25 |
|
3 |
8,21±0,26 |
29,54±2,36 |
166,02±10,69 |
1,15±0,36 |
|
|
6 |
8,32±0,41 |
29,21±2,17 |
170,31±12,58 |
1,00±0,28 |
|
|
9 |
9,01±0,26* |
29,31±3,16 |
182,36±14,01 |
1,01±0,14 |
|
|
Дегидратация |
12 |
14,36±0,71* |
62,10±4,21* |
218,31±16,57* |
2,71±0,23* |
|
15 |
17,01±0,66* |
65,21±4,12* |
217,32±20,41* |
2,51±0,17* |
|
|
18 |
18,95±1,12* |
69,21±5,01* |
215,20±20,17* |
2,69±0,31* |
|
|
21 |
23,01±2,00* |
70,21±3,17* |
213,31±20,01* |
2,74±0,24* |
|
|
24 |
25,10±1,98* |
79,00±5,28* |
214,36±19,99* |
2,62±0,21* |
|
|
Опытная группа |
|||||
|
Гидратация |
0 |
11,98±1,02 |
29,23±2,97 |
130,47±9,25 |
1,00±0,13 |
|
3 |
12,28±1,09 |
39,25±3,00* |
53,91±12,98* |
1,01±0,17 |
|
|
6 |
21,36±2,17* |
41,01±3,58* |
55,32±10,58* |
1,13±0,13 |
|
|
9 |
23,69±2,00* |
58,03±4,01* |
66,78±10,87* |
1,25±0,15 |
|
|
Дегидратация |
12 |
25,68±2,14* |
63,54±4,58* |
38,15±8,25* |
1,49±0,13* |
|
15 |
26,39±1,99* |
65,28±5,01* |
20,25±8,36* |
1,53±0,19** |
|
|
18 |
28,00±2,61* |
70,59±6,14* |
38,25±10,25* |
1,89±0,19* |
|
|
21 |
28,54±2,21* |
73,54±5,01* |
31,36±8,36* |
2,13±0,14* |
|
|
24 |
29,14±1,13* |
83,21±5,31* |
25,74±0,77* |
2,25±0,14* |
|
Примечание: * - р<0,05 - достоверность различий относительно 0 суток
Средняя исходная концентрация аргинин-вазотоцин в плазме у шести случайно выбранных птиц (0 сутки) составила 8,17±0,13. В период адаптации к высокосолевому рациону концентрация аргинин-вазотоцина в плазме крови птиц опытной группы возрастала.
К 12-м суткам эксперимента между всеми группами птиц наблюдались статистически значимые различия в уровне аргинин-вазотоцина в плазме крови, который демонстрировал прямую зависимость от уровня потребления хлорида натрия. У контрольных птиц изучаемый показатель повысился на 75,8%, у опытной группы птиц - 2,1 раза.
Можно предположить, что к 12-м суткам птицы были полностью адаптированы к рациону с повышенным содержанием соли и что осмоляльность плазмы является ключевым стимулом для секреции аргинина-вазотоцина. В данном случае просматривается корреляционная связь зависимости аргинина-вазотоцина от осмоляльности плазмы в условиях гидратации. Коэффициент корреляции в контрольной группе составил - 0,958, в опытной - 0,987.
Исходная концентрация альдостерона в плазме крови птиц опытной группы составляла 130,47±9,25 нг/л. На фоне потребления рационов с высоким содержанием хлорида натрия уровень альдостерона значительно снизился в течение 12 суток до 25,74± 0,77 нг/мл (р<0,05). В то же время, в контрольной группе уток, остававшейся на низкосолевом рационе, базальный уровень гормона сохранялся (табл. 2).
У дегидратированных птиц концентрация альдостерона демонстрировала слабую отрицательную корреляцию с осмоляльностью, натрия и хлора.
Исходная утренняя концентрация кортикостерона в плазме крови контрольных птиц (0 сутки) составила 1,10±0,25 нг/мл, в период обезвоживания наблюдалась общая тенденция к повышению концентрации кортикостерона до 2,62±0,21 нг/мл.
Наиболее быстрое и значительное увеличение кортикостерона было зарегистрировано во всех группах птиц, рост концентрации стал заметен уже на 2-е сутки дегидратации (12-е сутки эксперимента).
Через трое суток после прекращения доступа к воде концентрация аргинина-вазотоцина возросла в 2 раза у птиц контрольной группы и в 2 раза у птиц в группе с повышенным содержанием соли в рационе.
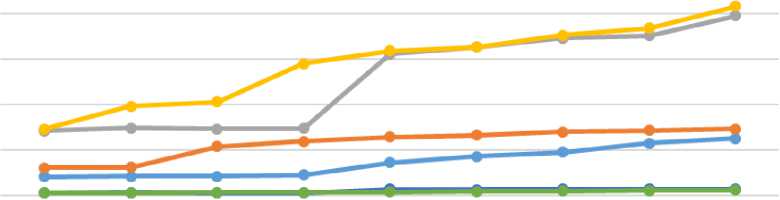
В последующие 9 суток значительного дальнейшего увеличения не наблюдали, что позволяет предположить, что секреция аргинина-вазотоцина достигла плато, несмотря на продолжающийся рост осмоляльности плазмы. Результаты исследований представлены на рисунке 1 и 2.
Обезвоживание вызвало повышение уровня кортикостерона во всех экспериментальных группах, что согласуется с данными других исследований. Хотя амплитуда ответа между группами значимо не различалась, анализ выявил различия в чувствительности: в контрольной группе зависимость кортикостерона от натрия была более выраженной (табл. 2).
Важно отметить, что увеличение кортикостерона в условиях дегидратации может быть не только осморегуляторной реакцией, но и следствием метаболического стресса, связанного со снижением потребления корма, которое также наблюдалось у наших птиц.
3 6 9 12 15 18 21 24
ДНИ
—•—контроль (A-В) —•—опыт (A-В) —«—контроль (П)—^-опыт (П) —^контроль (К) —^опыт (К)
Рисунок 1 - Влияние водной депривации на уровни аргинин-вазотоцин (А-В), (нг/л), пролактина (П), (нг/л) и кортикостерона (К), (нг/мл) у уток
Обезвоживание у птиц вызывает два ключевых физиологических сдвига: гиперосмоляльность и гиповолемию. Важно отметить, что выявленные различия в осмотической чувствительности секреции аргинина-вазотоцина между группами с разным потреблением соли не повлияли на конечную величину гормонального ответа. Во всех трех группах уровень аргинина-вазотоцина стабилизировался примерно на одном и том же значении (около 2529 нг/мл), несмотря на сохранявшуюся разницу в осмоляльности плазмы (на 11,6% между группами) и ее непрерывный рост на протяжении всего периода дегидратации. Этот факт позволяет предположить, что в условиях длительного стресса осмотические и объемные факторы способны поддерживать повышенную концентрацию аргинин-вазотоцин лишь до определенного предела, который, вероятно, соответствует максимальным возможностям его синтеза и высвобождения [6, 7].
Наблюдали четкую зависимость базового уровня пролактина от потребления хлорида натрия, которая проявлялась в виде ступенчатого увеличения гормона по мере роста содержания соли в рационе. Концентрация пролактина демонстрировала высокодостоверную положительную корреляцию с осмоляльностью плазмы, натрия и хлора.
В ответ на дегидратацию концентрация пролактина, демонстрировала резкое увеличение в 2-4 раза в течение первых двух суток. Однако в динамике пролактина и аргинина-вазотоцина выявлена существенная диссоциация. В отличие от аргинин-вазотоцина, который быстро достигал плато, концентрация пролактина продолжала неуклонно возрастать на протяжении всего периода обезвоживания во всех группах, причем наиболее значительный прирост наблюдался в опытной группе (рис. 1).
К концу периода дегидратации (дни 21-24) концентрация пролактина достигла исходного уровня во всех группах (80-85 нг/мл), что, вероятно, представляет физиологический максимум секреторного ответа на комбинированные осмотические и волюмические стимулы у уток. Можно предположить, что изначально более высокий осмотический статус и гиперволемия у птиц на высокосолевом рационе привели к снижению чувствительности осмо-регуляторного механизма, контролирующего секрецию пролактина [8].
Исходный уровень пролактина в плазме на 0-е сутки составлял 28-29 нг/мл. В период адаптации к рационам наблюдали увеличение пролактина у птиц, получавших высокое содержание хлорида натрия, причем наиболее выраженный рост отмечали у уток группы с повышенным содержанием соли в рационе. Так рост данного показателя к 24 суткам составил 2,4 раза.
Корреляционный анализ выявил значимую зависимость уровня пролактина от осмоляльности плазмы. Коэффициент корреляции в контрольной группе составил – 0,995, в опытной - 0,911.
В период обезвоживания наблюдали параллельный рост как осмоляльности плазмы, так и концентрации пролактина у птиц всех групп.
Положительная корреляция между концентрациями аргинина-вазотоцина и пролактина наблюдалась при анализе данных по группам: коэффициент корреляции для контрольной группы составил 0,957 для опытной группы – 0,932.
Несмотря на различия в динамике, средние объединенные значения кортикостерона между группами в период дегидратации у птиц с повышенным содержанием соли в рационе достигали статистической значимости. Однако сам факт повышения концентрации кортикостерона на фоне обезвоживания был статистически высокодостоверен.
Изменение содержания хлорида натрия в рационе и лишение организма доступа к воде, как известно, оказывают комплексное влияние на водно-солевой обмен, воздействуя как на объем внеклеточной жидкости (включая объем плазмы), так и на концентрацию электролитов. Это справедливо как для птиц, так и для млекопитающих [4]. Следовательно, изменения плазменных концентраций гормонов, зафиксированные в нашем исследовании, могут регулироваться как объемными, так и осмотическими стимулами.
В контексте домашней птицы, установлено, что десятикратное снижение содержания хлорида натрия в рационе приводило к уменьшению объема внеклеточной жидкости примерно на 10%, хотя объем плазмы при этом существенно не менялся. Это позволяет предположить, что в условиях нормальной гидратации объем внеклеточной (и, возможно, плазменной) жидкости был существенно ниже в контрольной группе по сравнению с группами, получавшими рацион с высоким содержанием соли. Кроме того, установлено, что водная депривация в течение 3-4 суток вызывает снижение объема плазмы на 16-20% у птиц со средним потреблением соли, что сопоставимо с условиями содержания наших птиц.
Обезвоживание, вызванное лишением воды, привело к резкому повышению осмоляльности, натрия и хлора уже ко второму дню во всех группах. На протяжении всего периода дегидратации наблюдался непрерывный рост этих показателей, причем наиболее медленные темпы увеличения были характерны для контрольной группы. Концентрация калия оставалась относительно стабильной.
Диапазон концентраций аргинин-вазотоцина в плазме у нормально гидратированных уток в нашем исследовании (8,179,01 нг/л), согласуется с данными, по концентрации базального уровеня антидиуретическо-го гормона, который в норме находится в нижней части этого диапазона [5].
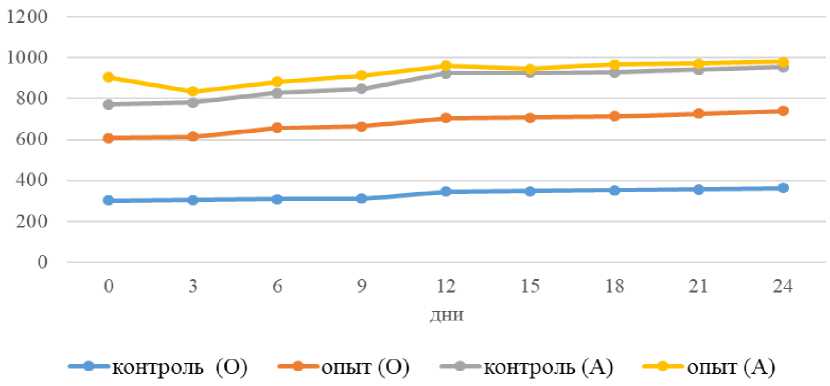
Рисунок 2 – Влияние водной депривации на осмомолярность (О), мОсм/л и уровень альдостерона (А), нг/л у уток
У млекопитающих установлено, что секреция аргинина-вазотоцина контролируется как осмотическими, так и объемными факторами.
Осморегуляторная система отличается более высокой чувствительностью, в то время как объемный фактор обладает большей мощностью. Кроме того, волемический статус оказывает обратное влияние на осмотическую чувствительность секреции аргинина-вазотоцина, для изменения которой требуется снижение объема крови не менее чем на 10% [3].
В нашем исследовании у уток с нормальной гидратацией наблюдалась положительная корреляционная зависимость между аргинин-вазотоцином и осмоляльностью плазмы. Наличие положительной зависимости аргинина-вазотоцина от осмоляльности у нормально гидратированных птиц, получавших разные рационы, указывает на доминирующую роль осмотического фактора над объемным, который должен был бы оказывать противоположное влияние. Однако это не исключает модулирующего эффекта волемии на осмотическую чувствительность.
Действительно, уровень потребления хлорида натрия и, как следствие, предполагаемый объем крови, по-видимому, влияли на чувствительность секреции аргинина-вазотоцина к осмоляльности, что особенно четко проявилось в условиях дегидратации.
Концентрация альдостерона в плазме, как известно, сильно зависит от потребления хлорида натрия у млекопитающих, рептилий и птиц. Наши данные, которые представлены на рисунке 2, полностью согласуются с этими данными [9].
У млекопитающих секреция альдостерона контролируется в основном ангиотензином II, калием и адренокортикотропным гормоном, причем натрий модулирует чувствительность надпочечников к этим стимулам. У птиц механизмы регуляции изучены хуже, хотя известно, что in vitro синтез стероидов в надпочечниках уток стимулируется дренокортикотроп-ным гормоном, калием и ангиотензином II.
Сочетание гиповолемии, высокой концентрации калия и низкой концентрации натрия в контрольной группе, по-видимому, привело к тому, что уровень альдостерона достиг своего максимума у нормогидратированных птиц. Об этом же свидетельствуют выявленные значимые корреляции: отрицательная с натрием, хлора и осмоляльностью, и положительная с калием.
То, что гиповолемия является мощным стимулом для секреции альдостерона, подтверждается нашим экспериментом с обезвоживанием. Несмотря на рост натрия, уровень альдостерона в контрольной группе не снизился, а даже несколько увеличился. Это можно объяснить компенсаторной реакцией на предполагаемое снижение объема крови, где эффект гиповолемии преодолевает влияние возросшего натрия [10].
Относительное увеличение альдостерона в опытной группе во время дегидратации было более значительным, однако достигнутые абсолютные значения составили лишь 50-80% от уровня контрольной группы. Это может быть следствием снижения чувствительности надпочечников к стимулам на фоне хронически высокого натрия.
Известно, что процедура взятия крови является стресс-фактором, влияющим на уровень кортикостерона. Однако в нашем эксперименте не было выявлено корреляции между продолжительностью забора крови и величиной кортикостерона у предварительно адаптированных птиц, что согласуется с данными о том, что обучение птиц процедурам снижает стрессовый ответ [9].
Несмотря на то что, имеются данные свидетельствующие в пользу осморегуляторной функции кортикостерона. Показано, что он частично восстанавливает электролитный баланс у адреналэктомированных уток, а его рецепторы обнаружены в почках. Кроме того, традиционное разделение кортикостероидов на минерало- и глюкокортикоиды может быть неприменимо к немлекопитающим, где один гормон часто выполняет обе функции.
Выводы. Исходные показатели водносолевого обмена у птиц на низкосолевом рационе были в пределах физиологической нормы. Девятидневная адаптация к рационам с разным содержанием хлорида натрия привела к формированию достоверных межгрупповых различий по осмоляльности плазмы, концентрациям натрия и хлора, которые соответствовали уровню потребления соли. Длительное обезвоживание вызвало прогрессирующий рост осмоляльности плазмы и концентраций ионов натрия и хлора во всех группах, причем у птиц на высокосолевом рационе эти параметры были достоверно выше. Концентрация калия демонстрировала меньшую динамику, оставаясь относительно стабильной в группе с высоким потреблением натрия в период дегидратации.
Гормональный ответ характеризовался значительным повышением уровня аргинин-вазотоцина, который находился в прямой зависимости от уровня потребления хлорида натрия и коррелировал с осмоляльностью плазмы в условиях гидратации. Однако в период обезвоживания, несмотря на продолжающийся рост осмоляльности, секреция аргинин-вазотоцина достигла плато, что указывает на предельную активацию системы. Уровень пролактина также достоверно повышался, демонстрируя тесную положительную корреляцию как с осмоляльностью, так и с концентрацией аргинин-вазотоцина. В отличие от них, концентрация альдостерона значительно снижалась на фоне высокого потребления хлорида натрия и демонстрировала слабую отрицательную корреляцию с осмоляльностью и электролитами. Концентрация кортикостерона, гормона стресса, закономерно увеличивалась в ответ на дегидратацию. Таким образом, длительное обезвоживание и уровень пищевого хлорида натрия вызывают комплексные и скоординированные изменения в осморегуляции и гормональном статусе домашней птицы.
