Влияние хронического психосоциального стресса на интенсивность дегенеративных изменений в сетчатке у крыс с различной стресс-резистентностью

Автор: Коршунова Динара Викторовна, Гиззатуллина Екатерина Александровна, Брындина Ирина Георгиевна
Рубрика: Фундаментальная медицина
Статья в выпуске: 5-4 т.16, 2014 года.
Бесплатный доступ
У 20 крыс моделировали хронический психосоциальный стресс в течение 10 и 18 дней, после чего изучали признаки апоптоза ганглиозных клеток сетчатки методом TUNEL и содержание церамида и сфингомиелина в тканях методом тонкослойной хроматографии. Проведено сравнение с контрольной группой (10 крыс). При стрессе обнаружены отдельные TUNEL-позитивные ядра ганглиозных клеток сетчатки, отсутствовавшие у контрольных животных. У стресс-неустойчивых крыс признаки дегенеративных изменений сетчатки (уменьшение количества ганглиозных клеток), как и повышение уровня церамида в структурах заднего сегмента глаз и плазме крови были более выражены, чем у стресс-устойчивых (в 5,9 раз и 4,3 раза соответственно). Полученные данные свидетельствуют о том, что психоэмоциональный стресс может способствовать развитию дегенеративных изменений в сетчатке, в большей степени у особей с низкой резистентностью к стрессу.
Апоптоз, ганглиозные клетки сетчатки, сфинголипиды, психосоциальный стресс, стресс-резистентность
Короткий адрес: https://sciup.org/148101909
IDR: 148101909 | УДК: 617.735:612.823:612.123:616
Influence of the chronic psychosocial stress on intensity of degenerate changes in retina at rats with various stress-resistance
At 20 rats it was modeled chronic psychosocial stress within 10 and 18 days, after then studied the signs of retina ganglionic cells apoptosis by TUNEL method and the maintenance of ceramide and sphingomyelin in tissues by method of thin layer chromatography. Comparison with control group (10 rats) is carried out. At a stress the separate TUNEL-the positive nucleus of retina ganglionic cells, which were absent at control animals are found. At a stress-unstable rats signs of degenerate changes of retina (reduction of ganglionic cells quantity), as well as increase the level of ceramide in structures of eyes posterior segment and blood plasma were more expressed, than at stress-steady (by 5,9 times and 4,3 times respectively). The obtained data testify that the psychoemotional stress can promote the development of degenerate changes in retina, in bigger degree at individuals with low resistance to stress.
Текст научной статьи Влияние хронического психосоциального стресса на интенсивность дегенеративных изменений в сетчатке у крыс с различной стресс-резистентностью
С постоянным ростом требований, предъявляемых социальной средой, все более значимым становится влияние стрессовых факторов на нервную систему и организм в целом. Показано, что психогенный стресс приводит к росту заболеваемости многими психосоматическими патологиями [2, 7]. Эмоциональный стресс является важным фактором снижения качества жизни и трудоспособности населения. В то же время клинические наблюдения свидетельствуют о существовании как людей, так и животных с различной пред-расположенностью к развитию нарушений в сходных стрессогенных ситуациях [5].
В настоящее время значимая роль в реализации стрессовых влияний на клеточном уровне отводится производным сфинголипидов: сфингозину, церамиду и др., которые считаются одними из ключевых сигнальных молекул [8]. Сфинголипиды широко изучаются в качестве внутри- и межклеточных регуляторов при различных видах клеточного стресса. Так, церамид, как вторичный мессенджер, вызывает остановку
роста и программированную гибель клеток. Показано участие церамида во всех известных механизмах инициации апоптоза [7]. Ранее нами было показано, что накопление церамида в различных тканях при стрессорных воздействиях существенно выше у животных с низкой устойчивостью к стрессу по сравнению с резистентными особями [1]. Апоптоз ганглиозных клеток сетчатки считается одним из ключевых звеньев развития глаукомной оптической нейропатии [6].
Цель исследования: оценить влияние психосоциального стресса на интенсивность дегенеративных изменений в сетчатке под действием эмоционального стресса.
Материалы и методы. Экспериментальные исследования проведены на 30 белых нелинейных крысах-самцах массой 180-220 г. с соблюдением всех регламентированных норм и правил этического обращения с лабораторными животными. Интенсивность дегенеративных изменений в сетчатке под действием эмоционального стресса оценивали гистологическими и иммуногистихимическими методами, также определяли содержание проапоптотического фактора церамида и его предшественника сфингомиелина в тканях заднего сегмента глаза и плазмы крови крыс методом тонкослойной хроматографии.
Для моделирования стресса 2-х месячных крыс на 2 месяца помещали в изолированные клетки при естественном световом режиме и свободном доступе к воде и пище. Перед помещением в условия изоляции, а также после нее животных тестировали согласно методу «открытого поля». По результатам тестирования были выделены 2 группы: стресс-устойчивые и стресс-неустойчивые (СУ и СН) крысы. После окончания периода изоляции животных ссаживали попарно с ежедневной сменой партнера. Проведено 2 серии опытов, в которых крысы подвергались воздействию 10-дневного (10 крыс) и 18-дневного (10 крыс) психосоциального стресса. 10 животных были интактными и послужили контролем. После окончания воздействий крыс выводили из эксперимента путем декапитации под тиопенталовым наркозом, глазные яблоки энуклеировали. Для гистологического и иммуногистохимического исследования глаза животных фиксировали в 10% нейтральном формалине 24 часа и использовали для приготовления парафиновых срезов согласно стандартному протоколу (Sennlaub F. et al., 2002). Срезы толщиной 5 мкм приклеивали на стекла с адгезивным покрытием (Silane-Prep Slides, Sigma). После депарафинирования для иммуногистохимического исследования срезы обрабатывали методом TUNEL (Terminal desoxynucleotidyl transferase – mediated desoxyuridine triphosphate (UTP) – nick end labeling). Использовали набор реактивов «Apoptag», Chemicon, США. В качестве субстрата для пероксидазы был применен диаминобензидин (DAB). Апоптотические ядра приобретали коричневое окрашивание. После реакции для дополнительного прокрашивания интактных ядер срезы обрабатывали 0,5% метиловым зеленым. Срезы для гистологического исследования окрашивали гематоксилином и эозином.
Для определения фракций сфинголипидов по экватору глазных яблок отсекали заднюю часть, из которой экстрагировали общие липиды смесью хлороформ/метанол 2:1 по Фолчу. Для разделения липидов на классы использовали метод восходящей хроматографии в тонком слое силикагеля по Boath et al. (2010). Использовали систему растворителей бутанол: ледяная уксусная кислота: вода (3:1:1) на пластинах фирмы «Merсk» с УФ индикатором (Германия). Пластины проявляли в парах кристаллического йода. Локализацию фракций на пластинах определяли с помощью «свидетелей» – стандартных хлороформенных растворов сфинголипидов фирмы Avanti polar lipids (CША). Расчет параметров и количественная оценка хроматограмм проводилась с помощью программы «Sorbfil TLC Videodensitometer».
В день получения материала забирали артериальную кровь, центрифугировали ее при 1000 об/мин в течение получаса для получения плазмы. Из 1 мл плазмы экстрагировали общие липиды смесью хлороформ/метанол 2:1 по Фол-чу и с помощью описанной выше методики восходящей тонкослойной хроматографии по Boath et al. (2010) разделяли липиды на классы. Количественную оценку хроматограмм (содер-жание в пятнах церамида и сфингомиелина) проводили так же, как описано выше. В другой порции плазмы (0,2 мл) определяли содержание 11-оксикортикостероидов по Резникову А.Г., 1980. Для оценки достоверности различий в группах использовали непараметрический критерий Манна-Уитни. Различия выборок считали статистически достоверными при р<0,05. Анализ зависимости между признаками проводили с помощью расчета r – коэффициент корреляции (r-критерия Спирмена).
Результаты и обсуждение. Иммуногистохимическое исследование сетчатки глаз позволило обнаружить у 2-х из 10-и животных опытной группы отдельные ТUNEL-положи-тельные ядра в ганглиозном слое сетчатки. В других слоях сетчатки апоптотических ядер выявлено не было. Таким образом, вовлеченными в апоптоз оказывались только ганглиозные клетки сетчатки, что может свидетельствовать об их большей уязвимости в сравнении с другими популяциями клеток ретины. Для определения интенсивности апоптоза в ходе гистологического исследования производили подсчет количества ядер ганглиозных клеток в срезах при увеличении ×400 от 300 мкм до 1300 мкм от края решетчатой пластинки (рис. 1). Как видно из диаграмм, количество ганглиозных клеток под действием зоосоциального стресса уменьшалось, что оказалось статистически значимым в группе СН крыс.
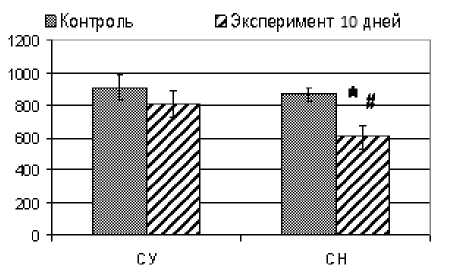
Рис. 1. Количество ядер ганглиозных клеток сетчатки сетчатки СУ и СН крыс при 10дневном зоосоциальном стрессе (*- p<0,05 в сравнении с контролем; # -p<0,05 – межгрупповые различия)
В контрольной серии содержание церамида и сфингомиелина у СН животных достоверно не отличалось от его количества в исследуемых тканях СУ крыс. Под действием 10 и 18 дневного психосоциального стресса как в заднем сегменте глаза, так и в плазме содержание церамида достоверно увеличилось в обеих группах в сравнении с контрольной, а количество сфингомиелина достоверно уменьшалось (табл. 1, 2). В группе СН животных увеличение оказалось более выраженным, чем в группе СУ крыс. Различия между типами животных в обеих сериях оказались достоверными (p<0,05).
Таблица 1. Сфинголипиды в заднем сегменте глаза у крыс с разной стресс-устойчивостью при психоциальном стрессе (в мкг на 1 мг)
|
Параметр |
Церамид |
Сфингомиелин |
||
|
СУ |
СН |
СУ |
СН |
|
|
контроль |
1,03±0,34 |
2,33±0,97 |
2,88±0,43 |
3,37±0,90 |
|
стресс (10 дней) |
4,47±0,90 * |
13,83±3,04 * # |
1,38±0,35 * |
0,50±0,10 * # |
|
стресс (18 дней) |
3,20±0,81 * |
6,10±1,30 * # |
1,77±0,20 * |
0,98±0,16 * # |
Примечание: здесь и далее *- p<0,05 в сравнении с контролем; # -p<0,05 – межгрупповые различия CУ – стрессоустойчивые, СН – стресснеустойчивые
Таблица 2. Сфинголипиды в плазме крови крыс с разной стресс-устойчивостью при психосоциальном стрессе (в мкг на 1 мл)
|
Параметр |
Церамид |
Сфингомиелин |
||
|
СУ |
СН |
СУ |
СН |
|
|
контроль |
0,034±0,007 |
0,041±0,009 |
0,021±0,004 |
0,016±0,0029 |
|
стресс (10 дней) |
0,072±0,01 * |
0,18±0,036 * # |
0,01±0,0018* |
0,004±0,0009* # |
|
стресс (18 дней) |
0,06±0,0087* |
0,132±0,027* # |
0,012±0,0011* |
0,008±0,0012* # |
Для выявления наличия или отсутствия связи между изменением содержания изучаемых фракций липидов в структурах заднего сегмента глаза и плазме артериальной крови был проведен корреляционный анализ. При сравнении содержания церамида в заднем сегменте глаза и плазме крови была выявлена сильная прямая зависимость, как у СУ, так и у СН животных (r=0,91 и r=0,88 соответственно, p<0,05). Это, по нашему мнению, может указывать на системный характер происходящего под действием стресса изменения обмена исследуемых сфинголипидов.
Аналогичная по силе и направленности зависимость наблюдалась и при сравнении содержания сфингомиелина в заднем сегменте глаз и плазме крови у обоих типов животных – СУ и СН (r=0,82 и r=0,87 соответственно, p<0,05), что может быть объяснено расходова-нием сфингомиелина в реакции образования церамида и также указывает на системность про-исходящих изменений. Сильная обратная зависимость с высоким уровнем достоверности была выявлена между содержанием церамида и сфингомиелина в тканях заднего сегмента глаз СУ и СН животных (r=- 0,91, p<0,05 и r=- 0,8, p<0,05 соответственно). Однако подобной достоверной связи не удалось обнаружить между содержанием церамида и сфингомиелина в плазме крови обоих типов животных, что, вероятно, может указывать на активность иных путей метаболизма церамида и сфингомиелина в разных тканях организма.
Это приводит к ослаблению корреляционных взаимосвязей на системном уровне.
Выводы: в эксперименте обнаружено нарастание содержания проапоптотического фактора – церамида, признаки дегенеративных изменений ганглиозного слоя сетчатки под действием зоосоциального стресса, зависящие от стресс-резистентности животных. Это позволяет предположить, что длительные повторные эпизоды стресса или тяжелое стрессовое воздействие может привести к более выраженным дегенеративным изменениям в сетчатке за счет накопления свободных радикалов, нейротоксических агентов (эксайтотоксичность) или за счет увеличения генерации церамида, способного активировать апоптоз клеток сетчатки [3, 5].
Исследование поддержано грантом ФГБУ «Фонд содействия развитию малых форм предприятий в научно-технической сфере» №0002918.
Список литературы Влияние хронического психосоциального стресса на интенсивность дегенеративных изменений в сетчатке у крыс с различной стресс-резистентностью
- Брындина, И.Г. Церамиды скелетных мышц, печени и легких грызунов при хроническом эмоциональном стрессе и моделированной невесомости/И.Г. Брындина, М.Р. Багаутдинов, Н.Н. Васильева и др.//Вестник Уральской медицинской академической науки. 2012. № 2(39). С.108-109.
- Судаков, К.В. Индивидуальность эмоционального стресса//Журнал неврологии и психологии. 2005. Т. 105, № 2. С. 4-12.
- Barak, A. Laser induces apoptosis and ceramide production in human retinal pigment epithelial cells/A. Barak, T. Goldkorn, L.S. Morse//Invest. Ophthalmol. Vis. Sci. 2005. Vol. 46. P. 2587-2591.
- Chida, Y. A bidirectional relationship between psychosocial factors and atopic disorders: A systematic review and meta-analysis/Y. Chida, M. Hamer, A. Steptoe//Psychosom. Med. 2008. Vol. 70. P. 102-116.
- Nora, P. Regulating survival and development in the retina: key roles for simple sphingolipids/P. Nora, E. Gisela, E. Carolina, O. German//J. Lipid Res. 2010. Vol. 51, № 6. P. 1247-1262
- Quigley, H.A. Neuronal death in glaucoma//Prog. Retin. Eye Res. 1999. Vol. 18. P. 39-57.
- Sanvicens, N. Ceramide is the key mediator of oxidative stress-induced apoptosis in retinal photoreceptor cells/N. Sanvicens, T. G. Cotter//J. Neurochem. 2006. Vol. 98. P. 1432-1444.
- Schenck, M. Ceramide: Physiological and pathophysiological aspects/M. Schenck et al.//Arch Biochem Biophys. 2007. Vol. 462. P. 171-175.
