Влияние различных видов ионизирующего излучения* и длительности культивирования на уровень генетической изменчивости сомаклонов сои

Автор: Фисенко П.П., Бабикова А.В., Ефремова О.С.
Статья в выпуске: 1 (142-143), 2010 года.
Бесплатный доступ
Методом маркирования межмикросателлитных последовательностей изучено генетическое разнооб-разие и взаимоотношения между растениями сорта Ходсон, мутантных популяций, полученных после обра-ботки семян различными видами излучения (Ходсон-L и Ходсон-γ) и их сомаклонами. Использование 12 праймеров позволило проследить вариабельность 183 ISSR-фрагментов, из них 89,61 % были полиморфными. Мутантные популяции Ходсон-L и Ходсон-γ характеризуются повышенным генетическим разнообразием по сравнению с популяциями сорта Ходсон. У сомаклонов, полученных из семядольных узлов растений мутант-ных популяций, уровень генетической изменчивости снижается и приближается к уровню изменчивости сорта Ходсон. Однако среди этих сомаклонов отмечены регенерантные линии, характеризующиеся более высоким содержанием белка, гистидина, линолевой и олеиновой кислот, а также в меньшей степени поражающиеся церкоспорозом и септориозом, чем растения их исходных форм. Длительность культивирования первичных эксплантов на питательной среде не влияет на уровень генетической изменчивости сомаклонов.
Соя, сомаклоны, уровень генетической изменчивости, виды ионизирующего излучения
Короткий адрес: https://sciup.org/142150939
IDR: 142150939
Influence of different kinds of ionizing radiation and cultivation time duration on genetic variability level of soybean somaclones
Genetic diversity and relations between plants of Hodson variety of mutant populations developed after seed treat-ment by different kinds of radiation (Hodson-L and Hodson-γ) and their somaclones were studied by means of inter-micro satellite sequence repeat (ISSR) markers. Usage of 12 ISSR-primers allowed to observe variability of 183 ISSR-fragments, 89.61 % of them were polymorphic. Mutant populations Hodson-L and Hodson-γ are characterized by richer genetic variabili-ty than populations of Hodson variety. Genetic variability level of somaclones developed from cotyledon's nodes of the mu-tant population plants decreases and becomes close to the level of Hodson variety variability. However, among these somaclones there were observed regenerant lines, which are characterized by the higher content of protein, histidin, linoleic and oleic acids, and also less damaged by Cercospora sojina and Septoria glycines, than plants of their parental forms. Culti-vation time of initial explants on medium does not influence on the level of the somaclones' genetic variability.
Текст научной статьи Влияние различных видов ионизирующего излучения* и длительности культивирования на уровень генетической изменчивости сомаклонов сои
Введение. Новая возможность расширения генетического разнообразия в селекции открылась с организацией биотехнологических методов исследований. Полученные на искусственных питательных средах в условиях in vitro растения-регенеранты, как правило, в той или иной степени отличаются от исходных форм и могут являться исходным материалом для традиционной селекции данной культуры, поскольку они представляют собой сомаклональные варианты [1].
Использование сомаклональной изменчивости в сочетании с отбором позволило О.А. Рожан-ской создать ценный селекционный материал сои, ярового рапса, нута, эспарцета, люцерны с признаками скороспелости, повышенной семенной и кормовой продуктивности, улучшенного химического состава, устойчивости к неблагоприятным гидротермическим условиям и патогенам [2].
В Казахстане группой ученых (С.В. Дидоренко, Ю.Г. Карягиным и Б.М. Жанысбаевым) сомакло-нальная изменчивость применяется как источник генетического разнообразия в создании новых форм сои [3].
В Приморском НИИСХ коллективом двух лабораторий: биотехнологии и селекции сои – с использованием метода культуры тканей создан первый в России сорт сои Приморская 81, который с 2004 г. районирован [4].
Важным моментом в технологии регенерации путем органогенеза являются условия, способствующие возникновению генетических изменений в рекомбинантах. По данным M.S. Wright и др. [5], дополнительные почки в пазухах семядольного узла сои закладываются de novo под влиянием 6-бензиламинопурина (БАП). R.A. Graybosch, M.E. Edge и X. Delannay [6] стимулировали побегообразование из вновь образовавшихся и ранее существовавших меристематических участков проводящей ткани семядольного узла на среде с БАП. Среди полученных линий исследователи наблюдали вариабильность по урожайности и другим признакам. A.H. Freytag с коллегами [7] изучали процесс регенерации растений сои из семядольного узла эксплантов эпикотиля. Авторы выявили генотипы, в потомстве регенерантов у которых отклонения от нормы появлялись чаще.
По мнению Рожанской [8], изоляция семядольных узлов на стадии развития проростка нарушает контролирующее влияние организма, приводит к неорганизованной пролиферации ткани и способствует возникновению генетических изменений у адвентивных почек, формирующихся в семядольных пазухах изолированных узлов.
Величины вариабельности признаков у регенерантов сои недостаточно высокие, поэтому с целью повышения генетического разнообразия мы использовали в качестве исходных эксплантов ткани мутантов. Существует много приёмов получения индуцированных мутаций. В основе их лежит воздействие на организм различными физическими и химическими факторами, называемыми мутагенами.
Действуя ими на растения, можно резко повысить их мутационную изменчивость. В селекционной работе используются любые виды ионизирующих излучений. Наиболее широко применяют рентгеновское, гамма- и нейтронное излучения. [9].
Наряду с этим существует вопрос о влиянии длительности культивирования на уровень генетической изменчивости сомаклонов сои. Как считают некоторые исследователи, уровень изменчивости со-маклонов повышается при более продолжительном культивировании ткани. Рядом авторов [10, 11] определено, что чем длительнее эксплант находится на питательной среде, тем больше вероятность получить регенеранты, отличающиеся от исходных форм.
Однако по данным M.S. Wright и др. [5], проводивших гистологический анализ пазух семядольного узла у исследуемых ими растений сои, через 6 дней после прорастания семян на среде с БАП базальные участки эпикотиля и семядолей, примыкающие к придаточным почкам, становятся меристематическими зонами. В эпидермальных и субэпидермальных тканях формируются проре-генеративные очаги, способные к морфогенезу, возникают новообразования – дополнительные адвентивные почки.
На основании вышеизложенных результатов и мнений ученых, нами была поставлена задача выяснить, оказывают ли влияние различные виды ионизирующего излучения и длительность культивирования на уровень генетической изменчивости сомаклонов сои.
Материалы и методы. Исследования проводились в лаборатории биотехнологии Приморского НИИСХ, в качестве исходных форм использовали сорт Ходсон и его мутанты. Для получения мутантов сухие семена обрабатывали в Институте цитологии и генетики СО РАН следующими электромагнитными излучениями: красным когерентным светом оптического квантового гелий-неонового генератора (лазера) с длиной волны 632,8 нм при плотности потока мощностью 0,08 мВт/см2 в течение 15 мин; γ-излучением кобальтовой пушки в дозе 50 грей.
В качестве первичных эксплантов использовали семядольные узлы стерильных микрорастений. Для получения последних зрелые семена стерилизовали в разделительной воронке концентрированной серной кислотой (H 2 SO 4 ) в течение 2-х мин, с последующей многократной отмывкой стерильной дистиллированной водой согласно методике, предложенной А.М. Смирновым, в изложении В.А. Тильбы [12].
Семядольные узлы культивировали на питательной среде 1/2MS + БАП (1,13 и 0,23 мг/л) до 60 дней. Адвентивные побеги, образовавшиеся de novo через 7-14 дней (R 0 -1) и через 31-60 дней (R 0 -2), снимали и помещали на среду 1/2MS+ИМК (0,5 мг/л), не содержащую БАП, для дальнейшего роста и развития.
Степень поражения (%) грибными патогенами: септориозом (Septoria glycines), церкоспорозом (Сercospora sojina), пероноспорозом (Peronospora manshurica) – определяли при искусственном заражении листовой поверхности на жестком инфекционном фоне совместно с сотрудниками лаборатории селекции сои ПримНИИСХ по методике ВИР [13] и согласно Международному классификатору [14].
Биохимический состав семян исходных форм и регенерантных линий проведен во ВНИИ сои на ИК-сканере Nir-42 по следующим показателям: аминокислоты (аргинин, валин, пролин, глютамин, лизин, гистидин, фенилаланин, тирозин, лейцин, изолейцин, аланин+глицин, треанин, серин, аспарагиновая кислота), жирные кислоты (пальмитиновая, стеариновая, олеиновая, линолевая, линоленовая), минеральные элементы (К, Са, Р, Мg), белок, масло. Оценка сомаклонов по химическому составу дана на основании классификатора [14].
Регенеранты третьего поколения выращивали в полевых условиях в соответствии с принятой для Приморского края агротехникой.
Статистическая обработка материала проведена методом дисперсионного анализа в изложении Б.А. Доспехова [15].
Для проведения генетического анализа все исследуемые образцы были разделены на три группы, каждая из которых была представлена растениями исходной формы и ее сомаклонами. Первая группа – сорт Ходсон и сомаклоны R 0 -1-690, R 0 -1-691, R 0 -1-699, R 0 -1-717, R 0 -1-722 и R 0 -2617. Вторая группа – популяция Ходсон- L , растения которой получены из облученных лазером семян, и сомаклоны R0-1-731, R0-2-616 и R0-2-623. Третья группа – популяция Ходсон-γ, растения которой получены из облученных γ-излучением семян, и сомаклоны R 0 -1-651, R 0 -1-688, R 0 -1-715 и R 0 -2-615.
Выделение ДНК и полимеразную цепную реакцию (ПЦР) проводили в БПИ ДВО РАН, как было описано нами ранее [16]. Для анализа полиморфизма межмикросателлитных последовательностей ДНК 52 образцов использовали 12 праймеров, комплементарных к микросателлитным повторам (табл. 1). При оценке электрофореграмм учитывали только четко видимые и воспроизводимые в повторных экспериментах фрагменты (ампликоны). Для каждого из праймеров были составлены бинарные матрицы, в которых присутствие или отсутствие в спектре фрагментов с одинаковыми молекулярными массами обозначали как "1" или "0". Разная интенсивность полос одинаковых по размеру ампликонов у сравниваемых образцов не учитывалась. Для определения длины фрагментов использовали маркер молекулярных масс – EcoRI + HindIII-рестрикты ДНК фага лямбда (Fermentas, Литва).
Таблица 1 – Праймеры, используемые в данной работе
|
Код праймера |
Нуклеотидная последовательность (5’–3’) |
Код праймера |
Нуклеотидная последовательность (5’–3’) |
|
пр812 |
(GA) 8 T |
прC1 |
(AGC) 6 T |
|
пр825 |
(AC) 8 T |
прC3 |
(AGC) 6 C |
|
пр840 |
(GA) 8 (CT) T |
прC4 |
(AGC)6G |
|
пр842 |
(GA) 8 (CT)G |
прC5 |
(TCG) 6 G |
|
пр888 |
(CGT) (ACT) (CGT) (CA) 7 |
прS1 |
(CA) 8 TG |
|
пр889 |
(AGT) (CGT) (AGT) (AC) 7 |
прS10 |
(GA) 8 TC |
Объединенная бинарная матрица была использована для расчета частот фрагментов, доли полиморфных локусов ( P ), генного разнообразия ( H ) и индекса Шеннона ( SI ) с помощью пакета программ POPGENE [17]. Для определения генетических расстояний Нея-Ли (D N ) и построения дендрограммы генетических взаимоотношений между отдельными растениями на основе значений D N посредством невзвешенного парно-группового метода кластерного анализа (UPGMA) с бутст-рэпными оценками степени надежности порядка ветвления (1000 реплик) использовали пакет программ TREECON [18, 19].
Результаты и обсуждения . В результате ISSR-анализа выявлено 183 фрагмента, из них 164 были полиморфными. Популяция Ходсон-γ характеризовалась наибольшим уровнем полиморфизма и значениями генного разнообразия и индекса Шеннона, чем все другие исследуемые популяции исходных форм (табл. 2).
Таблица 2 – Основные показатели генетической изменчивости исходных форм сои
|
Сорт/популяция |
Доля полиморфных локусов ( P, %) |
Генное разнообразие ( H ) |
Индекс Шеннона ( SI ) |
|
Ходсон |
12,57 |
0,041 |
0,062 |
|
Ходсон- L |
10,93 |
0,040 |
0,062 |
|
Ходсон-γ |
18,03 |
0,063 |
0,099 |
Генетические дистанции (DN) между парами анализируемых образцов (популяций) варьировали, достигая 10-кратного различия (табл. 3). Наименьшее значение D N (0,0642) отмечено между исходными популяциями Ходсон-γ и Ходсон- L , наибольшее (0,6043) – между сомаклоном R0-2616 популяции Ходсон- L и сомаклоном R0-1-691 сорта Ходсон.
Таблица 3 – Матрица значений генетических различий (D N ) между исследуемыми образцами сои, рассчитанных по 183 ISSR-фрагментам
|
Образец (популяция) |
Ходсон (n=10) |
R 0 -1690 (Ходсон) |
R 0 -1691 (Ходсон) |
R 0 -1699 (Ходсон) |
R 0 -1717 (Ходсон) |
R 0 -1722 (Ходсон) |
R 0 -2617 (Ходсон) |
Ходсон L (n=13) |
R 0 -1731 (Ход-сон-L) |
R 0 -2616 (Ход-сон-L) |
R 0 -2623 (Ход-сон-L) |
Ходсон γ (n=16) |
R 0 -1651 (Ход-сон-γ) |
R 0 -1688 (Ход-сон-γ) |
R 0 -1715 (Ход-сон-γ) |
R 0 -2615 (Ход-сон-γ) |
|
Ходсон (n=10) |
***** |
|||||||||||||||
|
R 0 -1-690 (Ходсон) |
0,179 |
***** |
||||||||||||||
|
R 0 -1-691 (Ходсон) |
0,545 |
0,447 |
***** |
|||||||||||||
|
R 0 -1-699 (Ходсон) |
0,337 |
0,261 |
0,381 |
***** |
||||||||||||
|
R 0 -1-717 (Ходсон) |
0,211 |
0,141 |
0,465 |
0,206 |
***** |
|||||||||||
|
R 0 -1-722 (Ходсон) |
0,165 |
0,092 |
0,414 |
0,247 |
0,116 |
***** |
||||||||||
|
R 0 -2-617 (Ходсон) |
0,199 |
0,141 |
0,518 |
0,319 |
0,166 |
0,153 |
***** |
|||||||||
|
Ходсон L (n=13) |
0,531 |
0,469 |
0,569 |
0,319 |
0,443 |
0,477 |
0,564 |
***** |
||||||||
|
R 0 -1-731 (Ходсон-L) |
0,210 |
0,172 |
0,509 |
0,254 |
0,134 |
0,160 |
0,186 |
0,439 |
***** |
|||||||
|
R 0 -2-616 (Ходсон-L) |
0,264 |
0,212 |
0,604 |
0,312 |
0,172 |
0,212 |
0,186 |
0,534 |
0,141 |
***** |
||||||
|
R 0 -2-623 (Ходсон-L) |
0,225 |
0,206 |
0,594 |
0,365 |
0,219 |
0,192 |
0,179 |
0,524 |
0,172 |
0,086 |
***** |
|||||
|
Ходсон γ (n=16) |
0,548 |
0,524 |
0,594 |
0,332 |
0,455 |
0,508 |
0,590 |
0,064 |
0,455 |
0,560 |
0,557 |
***** |
||||
|
R 0 -1-651 (Ходсон-γ) |
0,272 |
0,172 |
0,358 |
0,199 |
0,147 |
0,147 |
0,226 |
0,365 |
0,166 |
0,247 |
0,268 |
0,374 |
***** |
|||
|
R 0 -1-688 (Ходсон-γ) |
0,217 |
0,128 |
0,334 |
0,179 |
0,116 |
0,104 |
0,206 |
0,446 |
0,160 |
0,240 |
0,275 |
0,459 |
0,110 |
***** |
||
|
R 0 -1-715 (Ходсон-γ) |
0,295 |
0,186 |
0,456 |
0,268 |
0,147 |
0,1725 |
0,240 |
0,339 |
0,166 |
0,219 |
0,254 |
0,368 |
0,141 |
0,160 |
***** |
|
|
R 0 -2-615 (Ходсон-γ) |
0,257 |
0,172 |
0,509 |
0,297 |
0,199 |
0,199 |
0,212 |
0,536 |
0,166 |
0,166 |
0,226 |
0,565 |
0,261 |
0,212 |
0,247 |
***** |
Наибольшими генетическими отличиями от исходной формы характеризуются сомаклоны R 0 -1-691 и R 0 -1-699 в первой группе, R 0 -2-616 и R 0 -2-623 – во второй и R 0 -2-615 – в третьей (табл. 4). Однако уровень генетической изменчивости исследованных сомаклонов не зависит от длительности культивирования первичных эксплантов на питательной среде.
Таблица 4 – Значения генетических дистанций между сомаклонами и выборками их исходных форм и длительность культивирования сомаклонов in vitro
|
Сомаклон |
Длительность культивирования in vitro, сут. |
Генетические дистанции ( D N) |
||
|
Ходсон |
Ходсон- L |
Ходсон-γ |
||
|
R 0 -1-690 (Ходсон) |
7-14 |
0,1789 |
0,4695 |
0,5235 |
|
R 0 -1-691 (Ходсон) |
7-14 |
0,5451 |
0,5694 |
0,5937 |
|
R 0 -1-699 (Ходсон) |
7-14 |
0,3369 |
0,3188 |
0,3324 |
|
R 0 -1-717 (Ходсон) |
7-14 |
0,2108 |
0,4433 |
0,4547 |
|
R 0 -1-722 (Ходсон) |
7-14 |
0,1646 |
0,4772 |
0,5084 |
|
R 0 -2-617 (Ходсон) |
32 |
0,1993 |
0,5645 |
0,5904 |
|
R 0 -1-731 (Ходсон- L ) |
7-14 |
0,2098 |
0,4393 |
0,4550 |
|
R 0 -2-616 (Ходсон- L ) |
48 |
0,2643 |
0,5340 |
0,5596 |
|
R 0 -2-623 (Ходсон- L ) |
57 |
0,2249 |
0,5245 |
0,5571 |
|
R 0 -1-651 (Ходсон-γ) |
7-14 |
0,2722 |
0,3650 |
0,3735 |
|
R 0 -1-688 (Ходсон-γ) |
7-14 |
0,2173 |
0,4456 |
0,4589 |
|
R 0 -1-715 (Ходсон-γ) |
7-14 |
0,2952 |
0,3392 |
0,3678 |
|
R 0 -2-615 (Ходсон-γ) |
31 |
0,2566 |
0,5355 |
0,5646 |
Длительность культивирования семядольных узлов сорта Ходсон не влияет на генетическую изменчивость его сомаклонов. В то время, как для возникновения генетической дифференциации сомаклонов популяций Ходсон- L и Ходсон-γ от их исходных форм, вероятно, необходимо более длительное культивирование (см. табл. 4).
К третьему поколению из числа изучаемых нами сомаклонов были исключены стерильные, слабофертильные формы с нежизнеспособными семенами, а также непродуктивные и характеризующиеся другими отрицательными признаками (R 0 -1-691, R 0 -1-699, R 0 -1-690, R 0 -2-617, R 0 -1-651 и R 0 -1-688). Среди оставшихся сомаклонов по биохимическим показателям (содержание белка, гистидина, линолевой и олеиновой кислот) исходную форму превышали: сомаклон R617 первой группы ( D N 0,1993), все сома-клональные линии второй группы и регенерант R615 третьей группы ( D N 0,5646) (табл. 5).
Кроме того, сомаклон R623 второй группы ( D N 0,5245) обладал высоким уровнем устойчивости к церкоспорозу, а регенерант R715 третьей группы ( D N 0,3678) характеризовался высоким уровнем устойчивости к пероноспорозу и церкоспорозу.
Таблица 5 – Сомаклональные линии сои третьего поколения, имеющие преимущество перед исходными формами по некоторым признакам
|
Исходная форма (и.ф.), выделившаяся сомаклональ-ная линия (R) |
Продук-тив-ность, г/раст. |
Содержание в семенах масла, % |
Содержание кислоты, % от общего количества масла в семенах |
Содержание белка в семенах, % |
Содержание гистидина, от общего количества аминокислот, % |
Степень поражения патогенами листовой поверхности, % |
||||
|
лино левая |
линоленовая |
олеи новая |
септо-риоз |
церко-спо-роз |
перо-но-спороз |
|||||
|
Ходсон (и.ф.) |
7,6 |
20,2 |
51,5 |
4,9 |
6,5 |
38,5 |
8,3 |
62,5 |
68,8 |
82,5 |
|
R 617 |
9,5* |
20,5 |
52,6* |
3,7 |
10,4* |
38,4 |
10,2* |
43,8 |
52,5 |
57,5 |
|
Ходсон-L (и.ф.) |
7,5 |
21,2 |
51,9 |
4,8 |
4,5 |
37,4 |
10,7 |
50,0 |
57,5 |
70,0 |
|
R 616 |
6,3 |
21,6 |
52,3 |
5,0 |
4,8 |
39,3* |
11,9* |
41,3 |
29,8* |
56,3 |
|
R 623 |
6,1 |
20,0 |
52,4 |
4,4 |
8,6 |
39,2* |
10,3 |
35,0* |
24,8* |
57,5 |
|
R 731 |
8,2 |
21,5 |
51,9 |
5,4 |
4,3 |
37,6 |
11,9* |
41,3 |
37,5 |
71,3 |
|
Ходсон-γ (и.ф.) |
7,7 |
20,5 |
52,2 |
5,5 |
9,0 |
37,7 |
10,3 |
41,3 |
52,5 |
58,8 |
|
R615 |
6,6 |
20,0 |
52,7 |
3,4* |
10,7* |
38,1* |
9,6 |
40,0 |
47,5 |
58,8 |
|
R 715 |
6,9 |
20,3 |
51,7 |
4,7 |
6,5 |
38,1* |
10,9 |
42,5 |
33,8* |
50,0* |
Примечание: * – достоверно превосходит исходную форму на 5% -ном уровне
Следует отметить, что регенерантные формы, имеющие наибольшие генетические отличия от исходных форм, не всегда выделяются по биохимическим показателям, тогда как сомаклоны со средним (R715) и низким (R617) уровнем генетической изменчивости могут превышать исходную форму по селекционным признакам. В результате исследований в третьем поколении были выделены сомаклоны: R616, R623 и R617, которые рекомендованы селекционерам в качестве исходного материала для селекции сои в Приморском крае.
На рисунке представлена некорневая дендрограмма генетических взаимоотношений между исследуемыми образцами. Анализируемые растения распределились в два кластера: первый объединяет с высокой степенью достоверности (индекс бутстрепа 100 %) все растения популяций Ход-сон- L и Ходсон-γ, которые группируются соответственно их происхождению; второй объединяет все растения исходного сорта Ходсон (индекс бутстрепа 100 %) и все исследуемые сомаклоны, за исключением двух (R0-1-691 и R0-1-699). Эти два сомаклона сорта Ходсон имеют наибольшие генетические отличия как от исходной формы, так и от всех других.
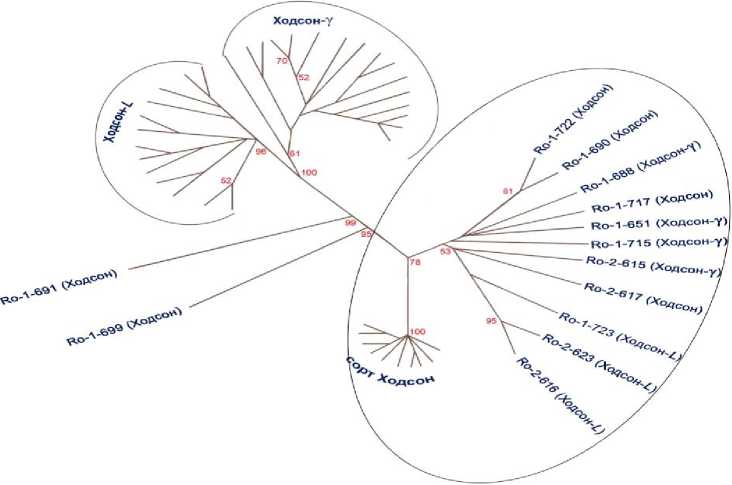
Рисунок – Некорневая дендрограмма генетических взаимоотношений между растениями сорта Ходсон, популяций Ходсон-γ и Ходсон- L и их сомаклонами.
Выводы. В результате проведенных исследований выявлено, что растения, выращенные из ткани обработанных различными видами облучения семян сорта Ходсон, генетически значительно отличаются от растений, выращенных из необработанных семян. Среди сомаклонов популяций Ходсон-γ и Ходсон- L выделены регенеранты, характеризующиеся повышенным содержанием белка, гистидина, линолевой и олеиновой кислот, а также более высокой устойчивостью к церкоспорозу и септориозу по сравнению с их исходными формами. Уровень генетической изменчивости сомаклонов не зависит от длительности культивирования первичных эксплантов на питательной среде.
Работа выполнена при финансовой поддержке гранта ДВО РАН №05-II-CХ-06-002C "Методы биотехнологии в селекции сои и риса".
