Зооархеологические и тафономические данные о средне- и верхнепалеолитических комплексах пещеры Бонди (Республика Грузия)
")
Автор: Йешурун Р., Монсель М.-Э., Плрдо Д., Пинхаси Р., Тушабрамишвили Н., Агапишвили Т., Лордкипанидзе Д.
Журнал: Археология, этнография и антропология Евразии @journal-aeae-ru
Рубрика: Палеоэкология. Каменный век
Статья в выпуске: 3 (59), 2014 года.
Бесплатный доступ
В статье представлены результаты детального зооархеологического и тафономического изучения материалов из пещеры Бонди в Западной Грузии. В ходе раскопок 2007-2010 гг. здесь выявлены средне- и верхнепалеолитические слои. В фа-унистическом комплексе преобладают кавказский тур (Capra caucasica) и европейский бизон (Bison cf. bonasus). Высокая доля костей животных со следами нарезок и ударов означает, что их накопление в основном связано с человеческой деятельностью. Характеристики комплекса позволяют предположить, что периоды обитания в пещере мобильных верхнепалеолитических охотников-собирателей были кратковременными. Состав фаунистических остатков свидетельствует о более открытых, по сравнению с голоценом, ландшафтах вокруг пещеры в верхнем палеолите.
Поздний плейстоцен, кавказ, охота, палеоэкология
Короткий адрес: https://sciup.org/14523056
IDR: 14523056 | УДК: 903
Текст научной статьи Зооархеологические и тафономические данные о средне- и верхнепалеолитических комплексах пещеры Бонди (Республика Грузия)
Изучение перехода от среднего палеолита к верхнему на Южном Кавказе важно для понимания природы замещения неандертальцев людьми современного антропологического типа в регионе, где могло происхо- дить их взаимодействие [Adler et al., 2008; Pinhasi et al., 2012; Moncel et al., in press]. Недавние исследования позволили получить большое количество данных о поведенческих характеристиках древних людей во время этого важного перехода, а также о темпах данного процесса. По всей видимости, и на Северном,
Археология, этнография и антропология Евразии 3 (59) 2014
и на Южном Кавказе неандертальцы и сопутствую- зультаты детального зооархеологического и тафоно- щие им среднепалеолитические индустрии исчезли за несколько тысячелетий до появления верхнепалеолитических индустрий, обычно рассматриваемых как созданные современными людьми (хотя антропологические находки, напрямую связанные с каменными артефактами, отсутствуют), что не позволяет говорить о непосредственном взаимодействии между двумя человеческими видами [Pinhasi et al., 2011; Pinhasi, Nioradze, Tushabramishvili et al., 2012]. Об этом разрыве свидетельствует и отсутствие преемственности в каменной технологии, а также другие аспекты материальной культуры, например, изделия из кости впервые появляются только в раннем верхнем палеолите [Adler et al., 2006; Pleurdeau et al., 2007; Bar-Yosef et al., 2011; Moncel et al., 2013]. В то же время недавние исследования показали, что на территории Западной Грузии системы жизнеобеспечения и до ставки обсидиана среднепалеолитическими неандертальцами были довольно похожи на те, которые практиковали антропологически современные люди в верхнем палеолите [Adler et al., 2006; Adler, Bar-Oz, 2009; Bar-Oz, Adler, 2005; Le Bourdonnec et al., 2012]. Обе популяции охотились почти исключительно на копытных, предпочитая взрослых животных, при этом развитие охотничьих навыков позволяло им перехватывать кавказских туров на путях их сезонных миграций. Способы транспортировки туш, их разделки и употребления в пищу были практически идентичны на позднем этапе среднего и в верхнем палеолите, что засвидетельствовано при изучении стратиграфических последовательностей ключевых памятников – пещер Ортвале Клде и
Дзудзуана [Adler et al., 2006; Adler, Bar-Oz, 2009; Bar-Oz, Adler, 2005; Бар-Оз и др., 2002; Bar-Oz, Belfer-Cohen, Meshveliani et al., 2008; Bar-Oz, Weissbrod, Gasparian et al., 2012; Moncel et al., 2013] (данные по Северному Кавказу см.: [Hoffecker, Cleghorn, 2000]). Эти наблюдения основываются на ограниченном наборе археологических источников, дополнительные средне- и верхнепалеолитические комплексы Западной Грузии могут позволить оценить, насколько отмеченные характеристики типичны для популяций неандертальцев и людей современного типа в данном регионе.
Недавние раскопки в пещере Бонди (Имеретия, Республика Грузия) дали фаунистические материалы, найденные в контексте слоев среднего и верхнего палеолита [Tushabramishvili et al., 2012]. Ре- мического исследования этого комплекса предоставляют дополнительные данные для изучения палеолита региона [Bar-Oz et al., 2012] и реконструкции систем охоты и жизнеобеспечения на южных границах хребтов Кавказа в позднем плейстоцене.
Стоянка и ее окружение
Пещера Бонди находится в бассейне рек Риони и Кви-рила в области Имерети и в Северо-Западной Грузии на высоте 477 м над ур. м. (рис. 1). Ландшафт этого района ныне всхолмленный и лесистый, изрезанный глубокими ущельями. Пещера расположена на южном склоне маленькой долины (рис. 2), находящейся примерно в 30 м выше р. Табагреби, в 2–4 км от пещерных стоянок Ортвале Клде и Дзудзуана.
В 2007–2010 гг. во фронтальной части пещеры, поблизости от современной капельной линии на площади 12 м2 была вскрыта толща отложений мощностью 2,8 м, включающая в себя восемь археологических горизонтов, разделенных слоями глыбовника и щебня, образовавшихся в результате крупных обвалов свода пещеры (рис. 3). Скальное основание пока не достигнуто [Tushabramishvili et al., 2012]. Раскопки выявили два основных культурных комплекса: среднего (финальный этап) и верхнего палеолита. Слой VII (среднепалеолитический, включает в себя мас- сивные пластины, удлиненные отщепы и свидетельства существования здесь леваллуазской технологии) датируется радиоуглеродным методом в интервале
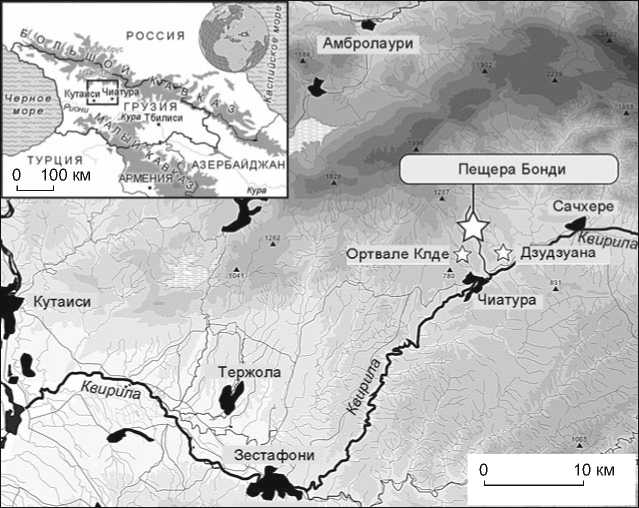
Рис. 1 . Расположение пещеры Бонди и других стоянок, упомянутых в тексте.

а

б
Рис. 2 . Вход в пещеру Бонди ( а ) и вид из нее ( б ). Июль 2011 г.
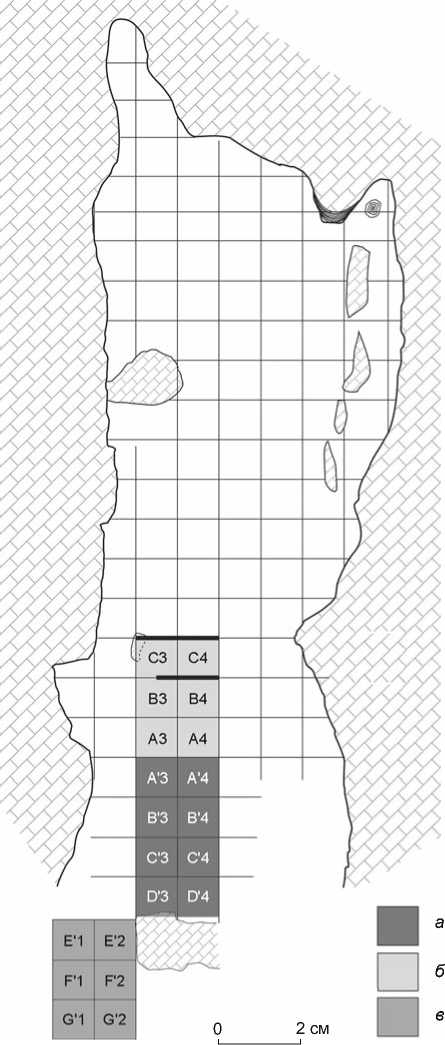
Рис. 3 . План раскопок в пещере Бонди. а – раскопки 2007 г.; б – 2009-го; в – 2010-го.
38 700–35 000 л.н. (калиброванные в системе Hulu значения – 43 000–40 000), слои V–III (верхнепалеолитические, на протяжении периода их формирования отмечается возрастание доли пластинок относительно пластин, в слое V найдена кость, отнесенная к роду Homo) – 24 600–14 000 л.н. (калиброванные в системе Hulu значения – 29 000–17 000) [Tushabramishvili et al., 2012]. Таким образом, слои верхнего палеолита в пещере Бонди могут быть частично одновременны с таковыми Дзудзуаны (пачки слоев D – B) и Ортвале Клде (слои 4d и 4c) [Moncel et al., in press].
Кости животных и каменные артефакты имеются во всех археологических горизонтах. В общей сложно сти найдено более 7 000 каменных артефактов, ок. 90 % из них – в слоях II, IV и V. В малочисленных комплексах нижних слоев VII и VIII присутствуют толстые удлиненные продукты ращепления, что резко отличает их от верхнепалеолитических, содержащих удлиненные тонкие пластины [Ibid.]. Свидетельства использования огня выявлены во всех слоях в виде обожженных кремней и микрофрагментов угля. Фаунистические материалы пещеры Бонди уже изучались ранее, однако исследования ограничивались лишь определением видов [Tushabramishvili et al., 2012].
Методы
Зоологические и тафономические данные получены согласно процедуре, описанной ранее [Bar-Oz, Adler, 2005; Bar-Oz et al., 2008]. Вся коллекция костей, собранная в процессе раскопок 2007–2010 гг., была просмотрена с целью поиска определимых частей скелета. Элементы, которые не могли быть однозначно отнесены к определенному слою или, по крайней мере, к периоду, не учитывались. Методика идентификации частей скелета была максимально расширенной и включала все определимые и пригодные для подсчета фрагменты костей. Таксономическое определение производилось с использованием сравнительной коллекции и остеологических каталогов [Schmid, 1972; Pales, Garcia, 1981]. Различение костей бычьих ( Bos или Bison ) осуществлялось по методике С. Дж. Олсена [Olsen, 1960], в разделении костей козьих ( Capra или Ovis ) мы следовали М.А. Цедеру и Х.А. Лапхэ-му [Zeder, Lapham, 2010]. Идентификация на уровне вида верифицировалась на основе фаунистических списков, разработанных А. Векуа и М. Бухсианидзе, которые использовали сравнительную коллекцию из Государственного музея Грузии в Тбилиси. Элементы скелетов, которые не могли быть отнесены к конкретному виду, группировались согласно классам размерности (например: мелкое млекопитающее, мелкое, среднее или крупное копытное).
С целью поиска различных следов воздействий поверхность костей осматривалась с помощью увеличительных линз с небольшой разрешающей способностью (×2,5). Фиксировались изменения, вызванные людьми (следы разделки, термического воздействия и преднамеренной фрагментации), животными (погрызы землеройных животных, следы дробления зубами хищника, воздействия желудочного сока), и постдепозиционные (выветривание, выщелачивание, дробления вследствие падения камней или ходьбы по костям, следы корневой системы растений, абразии) (по: [Behrensmeyer, 1978; Fisher, 1995]). Фрагменты длинных трубчатых костей, имевшие признаки слома в древности, изучались на предмет морфологии плоскостей изломов. Их очертания и углы учитывались с целью определения, были ли ко сти сломаны в сухом или свежем состоянии (детальное описание типов изломов см.: [Villa, Mahieu, 1991]). Также учитывалась степень целостности окружности длинных трубчатых костей (целая, больше или меньше половины), служившая в качестве индекса фрагментации (по: [Bunn, 1983]). Кроме того, оценка возраста основывалась на изменениях внешней поверхности длинных костей (степени срастания головок), данных о прорезывании и степени износа зубов нижней челюсти [Grant, 1982; Stiner, 1990].
Результаты
В результате раскопок 2007–2010 гг. в пещере Бонди была получена средняя по размерам коллекция костей животных, состоящая в основном из фрагментов длинных трубчатых костей средних и крупных копытных, хотя имеется некоторое количество целых костей и челюстей. Всего обнаружено ок. 5 900 фрагментов размером более 1 см, в т.ч. 12 идентифицированных экземпляров (NISP) в среднепалеолитических слоях и 277 – в верхнепалеолитических. Поверхность костей в основном хорошей сохранности.
В отложениях обоих периодов доминирующими таксонами являются кавказский тур ( Capra caucasica ) и бизон, скорее всего его европейская разновидность ( Bison cf. bonasus ) (табл. 1). Другие копытные представлены единичными образцами. К ним относятся благородный олень ( Cervus elaphus ) и ко суля ( Capreolus capreolus ). Остатки хищников присутствуют в отложениях обоих периодов. Идентифицированы волк ( Canis lupus ) и медведь ( Ursus sp.). Судя по относительно небольшому размеру костей, медведь представлен видом U. arctos , однако немногочисленность имеющихся в настоящее время замеров не позволяет утверждать это с определенностью (табл. 2). Значительную часть коллекции составляют кости (в основном фрагменты диафизов длинных костей), которые можно лишь распределить по размеру. Такая классификация соответствует видовому соотношению: подавляющее большинство фрагментов принадлежит крупным копытным (вероятно, бизонам) и средним (большинство здесь, видимо, составляют туры, меньшинство – благородные олени). Присутствие зубра ( Bos primigenius ) фиксируется на соседних стоянках [Bar-Oz et al., 2008], в отличие от комплекса пещеры Бонди, где все определимые кости жвачных демонстрируют морфологические характеристики бизонов.
Хотя коллекция из среднепалеолитических слоев очень мала (вероятно, потому что раскопками пока была вскрыта только верхняя часть отложений среднего палеолита), нам удалось выявить важное различие в таксономическом составе комплексов среднего и верхнего палеолита (рис. 4). Удельный вес остатков крупных хищников (волк и медведь) относительно доли костей копытных животных значительно больше в среднепалеолитических слоях (χ2 = 10,14, p = 0,001).
Таблица 1 . Таксономический состав коллекции
|
Таксон |
Средний палеолит |
Верхний палеолит |
||
|
NISP |
MNI |
NISP |
MNI |
|
|
Bison cf . bonasus |
2 |
Копытные 1 |
34 |
4 |
|
Крупные копытные |
– |
– |
66 |
– |
|
Capra caucasica |
1 |
1 |
38 |
3 |
|
Cervus elaphus |
1 |
1 |
2 |
1 |
|
Оленевые |
1 |
– |
1 |
– |
|
Средние копытные |
3 |
– |
102 |
– |
|
Capreolus capreolus |
– |
– |
1 |
1 |
|
Мелкие копытные |
– |
– |
10 |
– |
|
Ursus sp. |
2 |
Хищники 1 |
7 |
1 |
|
Canis lupus |
2 |
1 |
8 |
3 |
|
Средние хищники |
– |
– |
1 |
– |
|
Мелкие хищники |
– |
– |
4 |
– |
|
Средние птицы |
– |
Птицы – |
3 |
– |
Примечание : NISP – количество идентифицированных костей, MNI – минимальное число особей для взаимоисключающих видов.
Таблица 2 . Измерения костей
|
Номер образца |
Год раскопок |
Квадрат |
Подквадрат |
Глубина залегания, м |
Период |
Слой |
Кость |
Измерения, мм |
|
1 |
2 |
3 |
4 |
5 |
6 |
7 |
8 |
9 |
|
Ursus sp. |
||||||||
|
190 |
2010 |
B4 |
Ab |
255–265 |
ВП |
V, VI |
Верхний клык |
L > 99,7; B = 32,3 * |
|
192 |
2010 |
B4 |
C |
395–400 |
СП |
VIII |
Клык |
L > 47,1; B = 13,7 * |
|
88 |
2009 |
C4 |
B |
205–215 |
ВП |
V |
P 4 |
A – P = 12,7; ML = 8,7 |
|
194 |
2010 |
B4 |
A |
375–380 |
СП |
VIII |
Лучевая |
BD = 67 |
|
230 |
2007 |
73 |
ВП |
Фаланга 1 |
BP = 18,2; BD = 15,1; GL = 43,2; SD = 12,1 |
|||
|
Canis lupus |
||||||||
|
174 |
2010 |
B3 |
B |
370–375 |
СП |
VII |
P4 |
B = 10,4; GB = 14,1; L = 24,7 |
|
167 |
2010 |
A3 |
B |
390–395 |
ВП |
VI |
P 4 |
L = 16,6; B = 7,5 |
|
278 |
2010 |
D′2–E′2 |
360 |
» |
» |
L = 21,2; B = 8,2 |
||
|
218 |
2007 |
80 |
» |
M 1 |
L = 24; B = 9,8 |
|||
|
125 |
2009 |
C4 |
bd |
195–205 |
» |
V |
» |
L = 30,1; B = 11,7 |
|
276 |
2010 |
B′3 |
C |
255–265 |
» |
V (?) |
M 2 + M 1 |
L = 9,9; B = 7,3; L = 22,2; B = 8,9; L molarrow = 40 |
|
131 |
2009 |
C4 |
bd |
185–195 |
» |
V |
Фаланга 2 |
BP = 10,3; BD = 9,4; GL = 25; SD = 7 |
|
Cervus elaphus |
||||||||
|
195 |
2010 |
B4 |
A |
365–375 |
СП |
VII |
Пяточная |
BP = 21,9; GL = 98,3 |
Окончание табл. 2
|
1 |
2 |
3 |
4 |
5 |
6 |
7 |
8 |
9 |
|
Capra caucasica |
||||||||
|
142 |
2009 |
A3 |
D |
285–295 |
ВП |
V |
Плечевая |
BD = 53,5; HDH = 20,7 |
|
177 |
2010 |
B3 |
B |
305–315 |
» |
VI |
» |
BT = 47,9; HTH = 22,3 |
|
99 |
2009 |
B4 |
A |
255–265 |
» |
V |
Пяточная |
BP = 45 |
|
140 |
2009 |
A3 |
Cd |
275–285 |
» |
V |
Пясть |
BP = 34,5 |
|
208 |
2007 |
74 |
» |
Фаланга 2 |
BP = 16,7; BD = 13,5; GL = 33,9; SD = 12,2 |
|||
|
264 |
2010 |
A′4 |
D |
205–215 Bis |
» on cf . bonas |
V (?) us |
Фаланга 3 |
DLS = 46,7; MBS = 10,2 |
|
139 |
2009 |
A3 |
Cd |
275–285 |
» |
V |
Лопатка |
BP = 54,5; GLP = 81,7; LG = 70,1 |
|
222 |
2007 |
109 |
» |
Фаланга 2 |
BP = 42,2; BD = 35,9; GL = 52,5; SD = 34,4 |
|||
|
223 |
2007 |
94 |
» |
Фаланга 3 |
DLS = 73,6 |
|||
|
123 |
2009 |
C4 |
bd |
195–205 |
» |
V |
» |
DLS = 76,5; LD = 65,5; MBS = 20,9 |
Примечания : методика измерений и обозначения по: [Driesch, 1976]; СП – средний палеолит, ВП – верхний палеолит.
*Замеры сделаны на сочленении эмали и дентина.
Рис. 4 . Структура верхне- и среднепалеолитических комплексов.
а – копытные; б – крупные хищники.
Отложения верхнего палеолита были разделены на три части: нижнюю (слой VI), среднюю (IV, V) и верхнюю (I–III). На протяжении всей последовательности доля костных остатков оленей и крупных хищников всегда мала, во всех слоях доминируют кости тура и бизона (рис. 5). Однако удельный вес этих двух таксонов изменяется на протяжении времени: в нижней и средней части отложений преобладает первый, в верхней – второй (верхний : средний уровни – χ2 = 7,76, p = 0,051).
Из-за малого размера выборки тафономическое происхождение среднепалеолитических фаунистических остатков определить трудно. Здесь были выявлены одна кость со следами резки (пяточная кость оленя) и одна с погрызом хищника (фрагмент берцовой кости копытного животного среднего размера). Эти находки могут говорить о том, что в образовании данного комплекса участвовали как человек, так и хищники. Относительно тафономии верхнепалеолитических фаунистических материалов имеется значительно больше данных (табл. 3). Антропогенное происхождение остатков копытных следует из
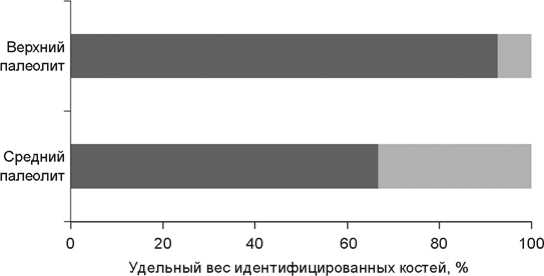

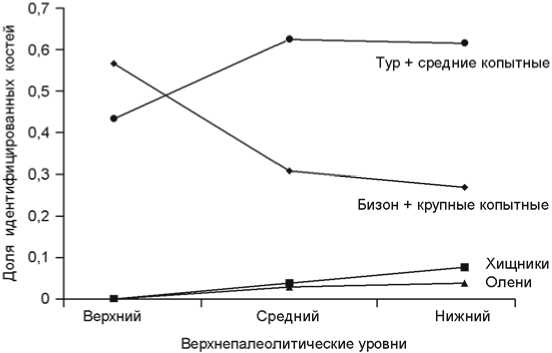
Рис. 5 . Тенденции в распределении частоты встречаемости видов в верхнепалеолитическом комплексе.
Таблица 3 . Данные об изменениях поверхности и фрагментации костей для каждой таксономической группы в верхнепалеолитическом комплексе
|
Признак |
Крупные хищники |
Мелкие |
Копытные |
|
|
средние |
крупные |
|||
|
NISP |
14 |
11 |
141 |
100 |
|
Следы воздействия огня |
0 |
0 |
1 (0,7 %) |
0 |
|
Излом свежий |
0 |
0 |
15 |
18 |
|
сухой |
0 |
3 |
11 |
6 |
|
промежуточный |
0 |
0 |
8 |
8 |
|
Целостность окружности труб- |
||||
|
чатой кости: |
||||
|
<50 % |
0 |
3 |
40 |
40 |
|
>50 % |
0 |
0 |
3 |
0 |
|
100 % |
2 |
2 |
3 |
0 |
|
Выветривание (стадии 3–5) |
0/7 * |
0/10 |
6 (4,8 %)/126 |
7 (9,1 %)/77 |
|
Нарезки |
0 |
0 |
11 (8,7 %) |
8 (10,4 %) |
|
След удара |
0/6 |
0/5 |
6 (9,7 %)/62 |
10 (20,8 %)/48 |
|
Погрыз хищника |
0 |
2 (20,0 %) |
5 (4,0 %) |
7 (9,1 %) |
|
Погрыз землероя |
0 |
0 |
0 |
0 |
|
Следы корней |
2 (28,6 %) |
3 (30,0 %) |
11 (8,7 %) |
12 (15,6 %) |
|
Следы топтания |
0 |
0 |
7 (5,6 %) |
7 (9,1 %) |
|
Абразия |
0 |
2 (20,0 %) |
7 (5,6 %) |
2 (2,6 %) |
*Здесь и далее после косой черты указано количество экземпляров, пригодных для определения данного признака.

0 2 cм
Рис. 6 . Образцы со следами воздействия человека и животного.
а – обломок большой берцовой кости (средняя часть) среднего копытного с признаками применения камня для ее дробления (широкая раковистая выемка, внутренний негатив скалывания отщепа, спиральный излом); б – фрагмент плечевой кости тура (№ 177), демонстрирующий погрызы хищника (зубчатые края, отметки зубов, узкая выемка без следов внутреннего скалывания отщепа).
высокого удельного веса костей с нарезками и следами ударов. Нарезки выявлены на костях из всех отделов скелетов туров и бизонов. Они отражают все этапы разделки туши (снятие шкуры, разделка, срезание мяса, чистка надкостницы). Характер излома костей свидетельствует об их дроблении с помощью камней. Большинство трубчатых костей конечностей сохраняют меньше половины своей изначальной окружности, что часто сопровождается «свежим» характером углов излома. Это может свидетельствовать о преднамеренном разбивании костей с целью извлечения их содержимого (рис. 6, а ). Странным кажется практически полное отсутствие обгоревших костей (один образец). На костных остатках хищных животных нет следов антропогенного воздействия. Активность хищников в пещере может быть засвидетельствована благодаря отметинам зубов на нескольких костях копытных (табл. 3, рис. 6, б ). Однако малое количество остатков хищников и умеренная частота встречаемости погрызов заставляет предположить, что их роль в образовании фаунистического комплекса верхнего палеолита была невелика. Влияние абиотических процессов разрушения (выветривание, утаптывание, воздействие корневой системы растений, абразии) на сохранность костей было незначительным (табл. 3).
Состав элементов скелета средних и крупных копытных в верхнепалеолитическом комплексе был реконструирован на основе подсчета минимального количества особей (MNI) для наиболее многочисленных определимых костей в каждом анатомическом отделе (табл. 4). Эти результаты могут быть использованы лишь с учетом поправки на сохранность костей, зависящую, в частности, и от плотности костных тканей. Фрагменты диафизов превосходят в количестве эпифизы одних и тех же костей конечностей средних и крупных копытных (соответственно 48 против 27 и 41 против 8). Это означает, что тафо- номические процессы по-разному воздействовали на элементы скелета, в зависимости от минеральной плотности костной ткани: губчатая эпифизов сохранялась хуже, чем кортикальная диафизов тех же самых костей.
Результаты анализа элементов скелета указывают на почти полную представленность анатомических частей средних копытных (тур), в то время как состав этих частей крупных копытных (бизон) выглядит более селективным (табл. 4, рис. 7): в основном нижние
Таблица 4 . Представленность элементов скелета средних и крупных копытных в верхнепалеолитическом комплексе
|
Элемент скелета |
Средние копытные |
Крупные копытные |
||
|
NISP |
MNI |
NISP |
MNI |
|
|
Череп |
7 |
3 |
4 |
2 |
|
Нижняя челюсть |
7 |
2 |
22 |
4 |
|
Шейный позвонок |
2 |
– |
– |
– |
|
Грудной/поясничный позвонок |
11 |
1 |
1 |
– |
|
Ребро |
20 |
– |
2 |
– |
|
Лопатка |
– |
– |
1 |
– |
|
Плечевая |
9 |
3 |
9 |
2 |
|
Пяточная |
7 |
2 |
5 |
1 |
|
Локтевая |
1 |
– |
1 |
– |
|
Кистевые |
5 |
– |
– |
– |
|
Таз |
4 |
– |
2 |
|
|
Бедренная |
5 |
– |
7 |
2 |
|
Большая берцовая |
9 |
3 |
18 |
5 |
|
Предплюсна |
7 |
– |
1 |
|
|
Плюсна |
24 |
2 |
6 |
1 |
|
Фаланга 1 |
11 |
2 |
6 |
– |
|
Фаланга 2 |
5 |
– |
3 |
– |
|
Фаланга 3 |
3 |
– |
2 |
– |
|
Сесамовидная |
2 |
– |
10 |
– |
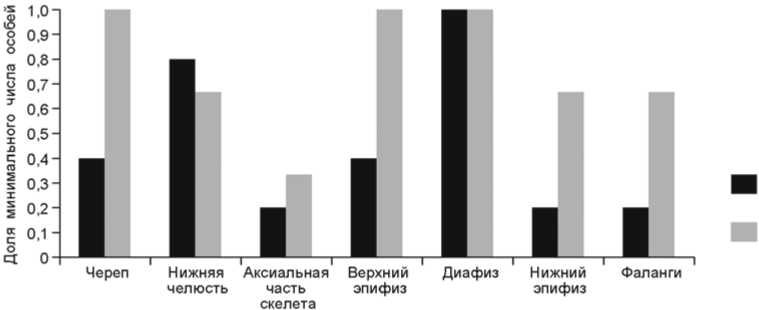
Рис. 7. Распределение элементов скелета крупных (а) и средних (б) копытных в верхнепалеолитическом комплексе.
челюсти и диафизы длинных ко стей конечно стей. Низкая представленность аксиальных частей скелетов в обоих размерных классах, вероятно, проистекает из различной плотности костей, что оказывало особое влияние на сохранность ребер и позвонков. Можно предположить, что туши туров транспортировались в полном или почти полном виде, в то время как значительно более крупные бизоны сначала разделывались на месте добычи, после чего самые выгодные в пищевом отношении части (содержащие много мяса или жира) переносились в пещеру.
Имеющаяся коллекция не позволяет реконструировать детальную картину возрастной структуры фаунистического комплекса. Исходя из анализа последовательности зубов dP4 – M3 со стадией износа в верхнепалеолитической коллекции, можно предположить, что в пищу были употреблены как минимум две взрослые особи тура. Судя по стадии прорезывания зубов и степени износа нижних P4, были добыты по крайней мере два молодых (нестертые зубы)
и два взрослых (стертые зубы) бизона. Степень сращения костей конечностей свидетельствует о превалировании взрослых особей туров и бизонов. Доля несиностозированных костных элементов составляет 14 % для Capra и 20 % для Bison (общее количество определимых индивидуальных элементов соответственно 35 и 10). Таким образом, на основании данных о зубах (стадия прорезывания и степень износа) и синостозировании ко стей можно предположить, что охотники добывали в основном взрослых туров. Относительно бизонов картина выглядит более расплывчатой и противоречивой из-за недостаточного размера выборки.
В пещере Бонди найдено пять обработанных костей (табл. 5, рис. 8). Все они происходят из верхнепалеолитических слоев, за исключением одного с неясным контекстом. Четыре предмета представляют собой шилья или их фрагменты. Одно изделие изготовлено из рога оленя, остальные – из трубчатых длинных костей жвачных животных. Все шилья, за исключени-
Таблица 5 . Образцы обработанных костей
|
Номер образца |
Местонахождение в раскопе |
Период |
Уровень |
Кость |
Таксон |
Длина, см |
Описание |
|
160 |
C3b, 245–255 м |
ВП |
V |
Конец рога |
Cervidae |
53 |
Нарезки – вероятно, результат обработки |
|
WB1 |
C4bd, 135–145 м |
» |
III–V |
ФДТК |
16 |
Продольные бороздки. Возможно, фрагмент шила |
|
|
WB2 |
E′2,580 м |
» |
Крупное копытное |
52 |
Острие шила |
||
|
WB3 |
B′4b, 225–235 м |
ВП |
» |
30 |
Почти целое шило |
||
|
WB4 |
B′4d, 210–225 м |
» |
» |
Среднее млекопитающее |
37 |
Продольные бороздки. Возможно, фрагмент шила |
Примечания : ВП – верхний палеолит, ФДТК – фрагмент длинной трубчатой кости.

а
Рис. 8 . Костяные орудия из пещеры Бонди.
а – WB3, шило; б – WB2, острие шила; в – WB4, предположительно фрагмент шила; г – № 160, обработанная верхушка рога; д – WB1, предположительно фрагмент шила.
ем одного, фрагментированы, ни одно из них не несет следов воздействия огня. Изделия из пещеры Бонди напоминают орудия этого типа из пещеры Дзудзуана [Bar-Yosef et al., 2011, fig. 5, 6], хотя менее вариабельны, вероятно, по причине меньшей коллекции.
Обсуждение
Пещера Бонди содержит средне- и верхнепалеолитические фаунистические комплексы, в которых доминируют тур и бизон. Это соответствует стратиграфическим последовательностям в Западной Грузии, особенно в расположенных поблизости пещерах Дзуд-зуана и Ортвале Клде [Bar-Oz, Adler, 2005; Adler et al., 2006; Bar-Oz et al., 2008; Adler, Bar-Oz, 2009; Bar-Yosef et al., 2011; Moncel et al., 2013]. Фаунистическая композиция также близка к той, которая была выявлена при изучении слоев позднего среднего палеолита на Северо-Западном Кавказе [Hoffecker, Cleghorn, 2000]. Наши выводы были построены в основном на верхнепалеолитической коллекции по причине ее достаточного для анализа размера. Эти фаунистические остатки свидетельствуют о том, что они появились в пещере в результате охоты, последующих транспортировки и разделки добытых животных для получения мяса и костного мозга. Способы транспортировки зависели от размеров туш. Копытные животные среднего размера (тур) приносились на стоянку целыми или почти целыми, а значительно более крупный бизон подвергался частичной разделке на месте добычи. Вполне возможно, что большие количества мяса бизона переносились на стоянку в виде отдельных кусков [Monahan, 1998]. Выброшенные остатки добычи поедались потом хищниками (особенно волком). Также они претерпевали разрушительное воздействие абиотических процессов, в результате чего сохранялись в основном кости, характеризующиеся более плотной структурой.
Вызывает интерес почти полное отсутствие обгоревших костей. На близлежащих палеолитических стоянках обнаружено много образцов со следами термического воздействия [Bar-Oz, Adler, 2005; Bar-Oz et al., 2008], что могло быть связано с использованием уже существовавших ко стных отбросов. Учитывая этот факт и то, что в пещере Бонди зафиксированы свидетельства деятельности хищников, а также умеренное количество костей и отдельных особей животных, мы склоняемся к выводу о коротких периодах обитания в пещере верхнепалеолитических людей, для которых была характерна высокая мобильность. Принимая во внимание большую долю остатков хищников в верхних среднепалеолитических слоях, можно предположить, что в среднем палеолите пещера использовалась еще менее интенсивно. Тем не менее, судя по составу элементов скелета животных и сле- дам антропогенного воздействия на костях, некоторые животные разделывались и употреблялись в пищу непосредственно на стоянке. Почти полная представленность частей тела тура позволяет предположить, что охотничьи угодья находились недалеко от пещеры. Наш вывод хорошо согласуется с особенностями среднепалеолитического каменного инвентаря, который очень немногочислен и представляет собой неполную редукционную последовательность. Это указывает на сезонное кратковременное заселение пещеры, когда приносились уже готовые орудия. Однако высокая насыщенность верхнепалеолитических слоев продуктами расщепления камня может свидетельствовать об утилизации местного кремня in situ и, соответственно, о другом характере заселения пещеры. Эти данные вносят свой вклад в дискуссию о типе использования окружающего ландшафта в среднем и верхнем палеолите популяциями района.
В конце верхнепалеолитической стратиграфической последовательности фиксируется большее пищевое использование бизона, нежели тура. Сокращение доли костных остатков бизонов на протяжении верхнего палеолита отмечалось также в пещере Дзудзуана, что воспринималось как отражение изменений в сезонности заселения стоянки [Bar-Oz et al., 2008; Bar-Yosef et al., 2011]. Для того чтобы определить, действительно ли эта тенденция связана с сезонностью, а также с изменениями в охотничьем инвентаре либо доступности тех или иных видов животных, требуются дополнительные исследования.
Два основных вида животных, представленные в фаунистических комплексах на протяжении всей стратиграфической последовательности пещеры Бонди, – кавказский тур и европейский бизон – прекрасно себя чувствуют в условиях гор, сочетающих как лесные, так и открытые ландшафты [Adler et al., 2006; Julien et al., 2012]. Позднеплейстоценовая макрофауна, реконструируемая по материалам памятника, может свидетельствовать о более низких высотных отметках и более открытых ландшафтах, нежели в голоцене, т.к. структура мезолитической (раннеголоценовой) фауны этого региона выглядит иначе. На близлежащей стоянке Котиас Клде в слоях, относящихся к мезолиту, ко стей тура и бизона нет, доминируют костные остатки бурого медведя, благородного оленя и косули, представлен также кабан (Sus scrofa) [Meshveliani et al., 2007; Бар-Oз и др., 2009]. Редкость или отсутствие трех последних видов в фаунистических комплексах пещеры Бонди и современных ей стоянок, вероятно, отражает условия значительно более открытых ландшафтов этого региона в поздний период среднего и в верхнем палеолите. По характеру фаунистического комплекса нами не отмечены заметные изменения окружающей среды в верхнем палеолите. Вероятно, ландшафтные условия в окрестностях пещеры и ее расположение на территории Западной Грузии способствовали поддержанию жизни здесь на протяжении большей части позднего плейстоцена, в отличие от местностей, альтитуда которых больше, или более удаленных от моря [Vekua, Lordkipanidze, 1998; Bar-Oz et al., 2012].
Мы благодарны А. Векуа и М. Бухсианидзе за помощь в таксономических определениях и Г. Бар-Озу за содействие в ходе исследований и замечания по рукописи этой статьи. Мы также признательны Министерству иностранных дел Франции и Национальному музею естественной истории (Париж) за финансовую поддержку в полевых работах и проведении анализов.
