Древесная растительность в пионерных позднеледниковых ландшафтах Прибалтики: результаты антракологического и фитолитного анализов

Автор: Дружинина О.А., Филиппова К.Г., Лазукова Л.И., Гольева А.А., Рудинская А.И., Бурко А.А., Сходнов И.Н.
Журнал: Общество. Среда. Развитие (Terra Humana) @terra-humana
Рубрика: Природная среда
Статья в выпуске: 4 (69), 2023 года.
Бесплатный доступ
Представлены результаты антракологического и фитолитного анализов двух позднеледниковых разрезов - Куликово и Утиное Болото (юго-восточная Прибалтика, Калининградская область). Оба разреза экспонируют отложения постледниковых водоемов, возникших в бёллинге - среднем дриасе, вероятно, при заполнении депрессий в рельефе талыми ледниковыми водами. Материалы исследования позволили рассмотреть вопрос о появлении и становлении древесной растительности на пост-ледниковых пространствах юго-восточной Прибалтики. Полученные результаты указывают на почти непрерывное локальное присутствие древесной (хвойной и лиственной) растительности на данной территории уже со среднего дриаса (не позднее ~13 980 кал.л.н.). Этот вывод хорошо согласуется с имеющимися палинологическими данными, свидетельствующими, что с ~13 840 кал.л.н. процентное содержание древесных пород не опускается ниже 60%, достигая 80% ~13 450 кал.л.н., к середине аллереда.
Антракология, древесная растительность, палинология, позднеледниковье, фитолиты, юго-восточная прибалтика
Короткий адрес: https://sciup.org/140305705
IDR: 140305705 | УДК: 551.89 | DOI: 10.53115/19975996_2023_04_282_287
Текст научной статьи Древесная растительность в пионерных позднеледниковых ландшафтах Прибалтики: результаты антракологического и фитолитного анализов
-
© Рудинская Анна Ивановна – младший научный сотрудник, Институт географии Российской Академии наук, Москва; e-mail: rudinskaya94@gmail.com
-
© Бурко Алексей Андреевич – младший научный сотрудник, НИЦ Прибалтийская археология, Калининград; e-mail: alesburk@mail.ru
-
© Сходнов Иван Николаевич – директор, НИЦ Прибалтийская археология, Калининград; e-mail: ivanskhodnov@gmail.com
Общество. Среда. Развитие № 4’2023
Палеогеографические исследования, проведенные в последнее десятилетие в Калининградской области (юго-восточная Прибалтика), свидетельствуют о том, что позднеледниковая природная среда на данной территории претерпела многочисленные изменения на финальном отрезке плейстоцена и в начале голоцена, связанные с таянием последнего Валдайского ледника и формированием современного облика ландшафтов [6; 8-10]. Вероятно, этот регион освободился от ледникового покрова в интервале 16-15 тыс. кал.л.н. [6]. Особенно динамичной в этот период была эволюция растительного покрова. На основании региональных спорово-пыльцевых данных, в целом, в позднеледниковом развитии растительности юго-восточной Прибалтики предполагаются следующие этапы: пионерные травяные
(полыневые, осоковые, злаки) и кустарничковые сообщества на освобожденных от ледника пространствах в раннем дриасе (16-14,7 тыс. л.н.); редколесья или «парковые» леса с сосной, березой и можжевельником в беллинге, а затем настоящие сосново-березовые леса в аллереде (14,712,9 тыс. л.н.); возможно, восстановление тундровых сообществ во время похолодания в среднем дриасе (14,1-14,0 тыс. л.н.), как, например, в более северных районах Прибалтики; и сокращение древесной растительности с увеличением открытых кустарничково-травянистых пространств в позднем дриасе (12,9-11,7 тыс. л.н.) [6; 8-10; 12; 16; 17].
При рассмотрении эволюции растительного покрова особого внимания заслуживает древесная растительность. Ее экологическая роль в формировании об-
* Исследование выполнено за счет гранта Российского научного фонда № 22-17-00113, project/22-17-00113.
лика позднеледниковых ландшафтов была значительна и заключалась во влиянии на микроклимат, формирование физико-химических свойств почвенного покрова, интенсивность поверхностной эрозии, а также на режим поверхностного стока. В эпоху позднего палеолита степень открытости ландшафта, наличие или отсутствие лесов, состав растительного покрова были одними из главных факторов, определяющих стратегии жизнеобеспечения древнего населения. В связи с этим понимание особенностей эволюции древесной растительности на конкретной территории важно для понимания существовавшей в прошлом палеоэкологической обстановки в целом. При этом одним из дискуссионных вопросов является доля древесных пород в общем составе растительности на освобожденных от ледника территориях, а также время их появления.
Самым обширным источником сведений о динамике древесного покрова являются палинологические данные. В Прибалтике пыльца древесных пород (Pinus, Betula, Salix, Picea) фиксируется уже в ранне -дриасовых отложениях. Большинство исследователей, однако, сходятся во мнении о заносном или переотложенном характере пыльцы древесных пород в данный период времени, учитывая ее единичные значения [6; 7]. Несмотря на то, что в отложениях бёллинга – среднего дриаса в Прибалтике количество пыльцы древесных пород возрастает (например, Pinus до 45%), особенности формирования спорово-пыльцевых спектров в условиях открытых пространств, существовавших в позднеледниковье, всё же заставляют относиться к подобным значениям с осторожностью [8]. Например, в современном спорово-пыльцевом спектре оз. Таймыр, которое в ботаническом отношении расположено в подзоне арктических тундр и где главнейшими компонентами зональных сообществ являются осоки и мхи, доминирует пыльца древесных и кустарников (66%). Среди древесных пород отмечена пыльца Betula sect. Albae, Pinus sylvestris, Pinus sibirica, Picea sp., Alnus sp., в группе кустарников и кустарничков – Betula sect. Fruticosae, Betula nana, Salix sp., Alnaster fruticosa. Подобный состав спектра сформировался преимущественно из заносной пыльцы и спор из более южных таежных районов, чему способствуют открытость окружающего озеро ландшафта (отсутствие древостоя, разреженность кустарничкового и травяного яруса, сомкнутого не более чем на 20-30%), большие размеры озера, сильные ветры [2]. Подоб- ные предпосылки формирования спектров могли существовать и в определенные периоды позднеледниковья. В результате, при использовании в палеореконструкциях только палинологической информации, может происходить пере- или недооценка роли древесного покрова в позднеледниковых ландшафтах. В то же время постепенно накапливаются другие категории палеоботанических данных, например, растительные макро- и микроостатков (древесина, кора, хвоя, фрагменты эпидермиса), которые служат достоверным дополнительным источником сведений в исследуемом вопросе и указывают на локальное присутствие древесных пород в Прибалтике с аллереда [16]. Внедряются также новые подходы и методы, к которым, в том числе, относятся антракологический и микробио-морфный (фитолитный) анализы.
Антракологический анализ основан на изучении углей в почве, озерных и торфяниковых отложениях с целью реконструкции пожаров естественного и антропогенного генезиса [14]. Важным для палеоботанических реконструкций является идентификация сгоревшего материала – древесины, листьев, игл хвойных, травы. Одним из направлений антраколо-гического анализа является также определение состава древесины, представленной в виде микроуглей.
Метод микробиоморфного анализа заключается в изучении комплекса микробиоморф – фитолитов, растительного детрита, кутикулярных слепков растений - и предоставляет широкие возможности для палеоэкологического анализа [4; 5]. Одна из категорий микробиоморф - фитолиты – являются кремниевыми слепками растительных клеток, формируемых растениями, произраставшими или использовавшимися на определенной территории. Индекационные формы фитолитов для лиственных древесных пород умеренного пояса к настоящему моменту не выявлены, однако они хорошо известны для хвойных пород. В отличие от органики, фитолиты устойчивы к разрушению и могут сохранятся в почве или в озерных осадках миллионы лет. Серьезным преимуществом фитолитов по сравнению с пыльцой является их низкая мобильность, поэтому фитолит-ный анализ способен предоставить сведения о растительном покрове локального уровня.
В 2022-2023 годах в Калининградской области начато изучение двух позднеледниковых разрезов – Куликово и Утиное Болото. Оба объекта экспонируют отложе-
Среда обитания
ния постледниковых водоемов, возникших в бёллинге, вероятно, при заполнении депрессий в рельефе талыми ледниковыми водами. В данной статье рассматриваются первые результаты антракологического и микробиоморфного исследования разрезов и их значение для решения вопроса о присутствии древесных пород в позднеледниковых ландшафтах.
Материалы и методы
Объекты исследований и географическая характеристика региона. Разрез Куликово расположен на западе Калининградской области, в береговом обрыве Самбийского полуострова (54°56'13" N, 20°21'30" E); разрез Утиное Болото - в восточной, внутриматериковой части территории (54°46’58" N, 22°25’23" E). В геоморфологическом отношении поверхность региона представляет собой чередование обширных равнинных и низменных пространств с отдельными холмисто-грядовыми возвышенностями [3]. Современный рельеф сформировался в результате непосредственной деятельности ледников последнего Валдайского оледенения, а также лимно- и флювиогляциальных процессов. Самбийский полуостров отличается довольно сложными сочетаниями форм: здесь природные комплексы холмистого, холмисто-грядового конечно-моренного и камового рельефа чередуются с плоскими и слабоволнистыми моренными и озерно-ледниковыми равнинами. На востоке области обширную террито рию занимает Шешупская озерно-ледниковая равнина. Она образовалась на месте приледникового водоема, сложена мелкозернистыми песками, супесями, глинами. В пределах равнины на фоне выровненной поверхности встречаются массивы и гряды моренных холмов, многочисленны камы и озы. В данной части Прибалтики под луговой, болотной и лесной растительностью (светлыми сосновыми, еловыми, широколиственными лесами, а также черноо-льшанниками) формируются различные типы почв - от болотных низинных перегнойно-торфяных до бурых лесных. Климат характеризуется избыточным увлажнением, мягкой зимой со средней температурой января -4,5°С и не жарким летом со средней температурой июля +17,5 °C.
Методы исследований . В результате бурения в 2022-2023 гг. на разрезах Куликово и Утиное Болото были отобраны керны мощностью 1,92 м и 1,89 м соответственно. Для обоих разрезов получены АМС 14С датировки, составляющие геохронологическую основу исследования (табл. 1, 2). Антракологический анализ включал подсчет и идентификацию макроскопических (линейные размеры >100 мкм) частиц угля в образцах [14]. Для этого проводилась обработка грунта 9% раствором перекиси водорода H 2 O 2 и промывание осадка на сите с диаметром ячейки 0,1 мм. Подсчет частиц макроуглей был выполнен на бинокулярном микроскопе MOTIC SMZ-168 при 20-кратном увеличении. Классифи-
Таблица 1
Результаты АМС 14С датирования разреза Куликово
Общество. Среда. Развитие № 4’2023
|
№ п.п. |
Глубина, см |
Возраст, С14 |
Возраст, кал.л.н. усредн. |
Материал |
|
1. |
192 |
12 200 ± 60 |
14 038 ± 160 |
Гиттия |
|
2. |
186 |
11 980 ± 80 |
13 957 ± 140 |
Дерево |
|
3. |
163 |
11 790 ± 60 |
13 693 ± 130 |
Дерево |
|
4. |
106 |
11 060 ± 60 |
13 102 ± 160 |
Дерево |
|
5. |
45 |
10 940 ± 60 |
12 773 ± 240 |
Дерево |
|
Таблица 2 Результаты АМС 14С датирования разреза Утиное Болото |
||||
|
№ п.п. |
Глубина, см |
Возраст, С14 |
Возраст, кал.л.н. усредн. |
Материал |
|
1. |
146 |
11 530 ± 60 |
13 505 – 13 295 (94,4 %) |
Гиттия |
|
2. |
120 |
11 190 ± 70 |
13 190 – 12 920 (91,0%) |
Гиттия |
|
3. |
100 |
9 135 ± 45 |
10 420 – 10 215 (92,0%) |
Дерево |
|
4. |
39 |
7 230 ± 45 |
8 175 – 7 960 (95,4%) |
Торф |
|
5. |
17 |
5 495 ± 40 |
6 245 – 6 200 (74,9 %) |
Торф |
кация углей по морфотипам и определение сгоревшего материала проводились согласно C.Mustaphi и M.Pisaric [14]. Исследование объекта Куликово включало 67 образцов, Утиное Болото - 55 образцов грунта.
К настоящему моменту микробиомор-фный анализ выполнен только для одного из объектов - разреза Куликово (60 образцов). Методика включала обработку образца 30 % перекисью водорода при комнатной температуре для разрушения органо-минеральных соединений и диспергации породы [4]. Затем высушенные образцы просматривались при помощи электронного микроскопа JEOL 6610LV и фотографировались. Морфологическое описание фитолитов составлялось согласно требованиям Международного кода по фитолитной номенклатуре [11], а их биоценотическая характеристика дана по А. Гольевой [13].
Результаты и обсуждение
Антракологический анализ . Из 67 исследованных образцов разреза Куликово уголь присутствует в 51. Обнаружены 22 морфо-типа углей, среди которых идентифицируются древесина, листья деревьев, листья злаков, хвоя, корни и другие растительные карбонизированные макроостатки. Морфо-типы, ассоциируемые с древесной растительностью, впервые фиксируются в слое, датируемом 13 987 ± 140 кал.л.н. (средний дриас), и обнаружены в 38 образцах, в целом, по всей толще разреза. Выделяются несколько коротких интервалов, где древесный материал не обнаружен; среди них самый продолжительный соответствует 13 095 - 13 230 кал.л.н., что совпадает с холодным событием GI-1b [15].
Исследование разреза Утиное Болото показало, что уголь содержится в 19 образцах из 25, соотносимых с позднелед-никовьем. Обнаружены 19 морфотипов; среди ботанических микроостатков идентифицируются аналогичные разрезу Куликово: древесина, листья деревьев, листья злаков и другие. Появление палеоботанических свидетельств древесных пород впервые фиксируется в слое, датируемом 14 400 ± 800 кал.л.н. В разрезе только 4 образца из 19 не содержат идентификаторов древесной растительности.
Микробиоморфный анализ. В образцах объекта Куликово обнаружены 6 индикационных форм фитолитов. В подавляющем большинстве это морфотипы, характерные для двудольных и ряда однодольных трав (разнотравье), включая злаки. Встречены формы, являющиеся диагностом тростника. Фитолиты, идентифицирующие иглы хвойных деревьев, найдены в 6 образцах в разных частях профиля – от самого низа до верха разреза, что охватывает интервал от среднего до позднего дриаса. Впервые данный морфотип встречается в слое, датируемом 13 980 ± 140 кал.л.н.
Позднеледниковая растительность юго-восточной Прибалтики в свете палинологических данных. Прежде всего, необходимо отметить, что позднеледниковая растительность на значительных пространствах Восточно-Европейской равнины обладала сходными чертами [1]. Она характеризовалась обилием степных и луговых трав (Cyperaceae, Poaceae, Artemisia, Fagopyrum sp., Valeriana sp. и др.), ксерофитов, толерантных к низкой зимней температуре, и гелиофитов ( Ephedra, Helianthemum ), а также растений, характерных для участков с нарушенными грунтами ( Artemisia, Amaranthus sp., Chenopodium album ). В следующую группу входили растения болот, влажных лугов и прибрежных местообитаний ( Equisetum, Polygonum ) и водные растения ( Myriophyllum, Nymphaea sp.). В составе флоры были также травянистые растения, связанные с бореальными лесными сообществами, такие как плауны и папоротники ( Lycopodium, Pteridium aquilinum ), арктоальпийские микротермные растения ( Betula nana, Selaginella selaginoides, Thalictrum ). Древесная растительность была представлена, прежде всего, деревьями с широкой экологической амплитудой ( Pinus sylvestris, Picea abies, Betula humilis ).
Всё описанное разнообразие видов растений в полной мере представлено в спорово-пыльцевом спектре разреза Куликово, имеющего самые ранние из изученных на сегодняшний день данных о составе позднеледниковой растительности юго-восточной Прибалтики [8]. Нижняя часть разреза соответствует интервалу 13 900 -13 400 кал.л.н. и позволяет проследить начальные этапы в становлении древесного покрова (рис. 1).
Слои разреза, датируемые 13 900 ± 160 кал.л.н., содержат пыльцу Pinus (45%), Betula sect. Albae (26%), Picea (3%), Salix (5%). В целом, на данном этапе вклад древесной растительности варьирует от 40 до 60%. Преобладают две пионерные породы – Pinus и Betula . Широкая представленность березы как пионерной породы связана с ее свето-любивостью, продуктивностью, с коротким вегетационным периодом. На быстрое распространение березы во время потеплений позднеледниковья могли влиять близость ее рефугиумов и скорость миграции. Сосна, имеющая тенденцию вытеснять березу, об-
Среда обитания
Общество. Среда. Развитие № 4’2023
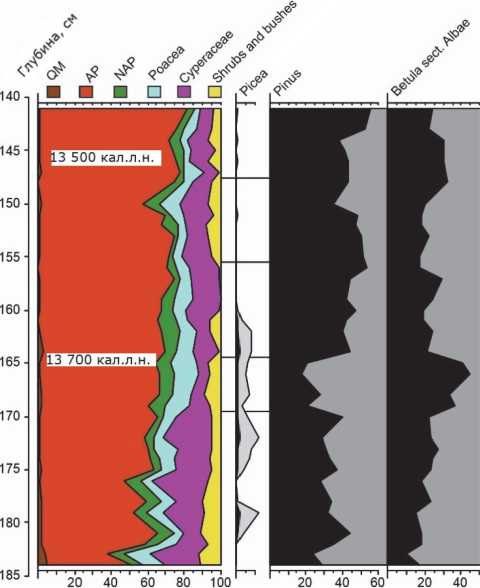
Рис. 1. Процентное содержание древесной растительности и основных древесных пород в палинологическом спектре разреза Куликово в интервале 13 900–13 400 кал.н.л. [8].
ладает более длительным вегетационным периодом, но не требовательная к климатическим и почвенно-грунтовым условиям.
В интервале 13 760–13 700 ± 140 кал.л.н. происходит изменение процентного состава древесных пород: содержание пыльцы сосны падает до 20%, в то время как березы возрастает до 45% при общем вкладе древесных пород, приближающемся к 70%.
Далее период аллерёда до 13 560 ± 160 кал.л.н. характеризуется колебаниями процентного соотношения двух главных древесных пород, при общем, очень постепенном, росте содержания древесной растительности до 75%. Количество пыльцы сосны варьирует между 45% и 55%, березы между 18% и 30%. Практически исчезает из спорово-пыльцевого спектра ель. Резкое сокращение древесных пород в растительном покрове (до 60%) наблюдается около 13 560 ± 160 кал.л.н. (возможно, как реакция на холодное событие GI-1c2 [15]), однако затем их доля возрастает и к 13 450 ± 160 кал.л.н. превышает 80%. Вклад отдельных древесных пород в это значение составляет Pinus (55%), Betula sect. Albae (25%), Picea (1,5–2%). В позднеледниковом спорово-пыльцевом спектре Куликово постоянно присутствуют также такие породы как Salix (до 5%), Alnus (до 5%), Corylus (до 3 %).
Заключение
Итак, согласно палинологическим данным, примерно с 13 900 ± 160 кал.л.н. начинается постепенный рост процентного содержания древесной растительности. С начала аллереда, 13 840 ± 140 кал.л.н. это значение не опускается ниже 60%, достигая 80% около 13 450 ± 170 кал.л.н., к середине периода. Палинологические данные хорошо согласуются с результатами антракологическо-го и микробиоморфного анализа, которые, дополняя друг друга, указывают на почти непрерывное локальное присутствие древесной (хвойной и лиственной) растительности, уже со среднего дриаса (не позднее ~13 980 кал.л.н.). Вероятно, и имеющиеся результаты спорово-пыльцевого анализа отражают эту картину, даже с поправкой на некоторое количество заносной пыльцы.
В заключение можно отметить, что ан-тракологический и фитолитный анализы могут служить как независимыми методами, так и быть важным дополнением к палинологическому анализу в реконструкции позднеледниковой растительности.
Список литературы Древесная растительность в пионерных позднеледниковых ландшафтах Прибалтики: результаты антракологического и фитолитного анализов
- Борисова О.К. Ландшафтно-климатические условия в центральной части Восточно-Европейской равнины в последние 22 тысячи лет (реконструкция по палеоботаническим данным) // Водные ресурсы. Т. 48. – 2021, № 6 – С. 664–675.
- База данных «Палеоклимат и изменения уровня моря в Арктике и Антарктике». – Интернет-ресурс. Режим доступа: paleopolar.aari.ru/index.php/uroven/arktika/20-ozero-tajmyr (06.11.2023)
- Географический атлас Калининградской области / Гл. ред. В.В. Орленок. – Калининград: Издательство КГУ, ЦНИТ, 2002. – 276 с.
- Гольева А.А. Микробиоморфные комплексы природных и антропогенных ландшафтов: Генезис, география, информационная роль. – М.: Издательство ЛКИ, 2008. – 240 с.
- Дружинина О.А. О перспективах применения метода фитолитного анализа в палеолимнологии // Естественные и технические науки. – 2020, №3. – С. 139–142.
- Дружинина О.А. Взаимодействие природы и общества в междуречье Немана и Вислы на рубеже плейстоцена и голоцена / Дисс. … канд. геогр. наук. – Калининград, 2005. – 160 с.
- Юспина Л.Ф. Палеогеография Балтийского моря: По данным палинологии верхнечетвертичной осадочной толщи / Дисс. … канд. геогр. наук. – Калининград, 2001. – 136 с.
- Druzhinina O., Rudinskaya A., Filippova K., Lazukova L., Lavrova N., Zharov A., Skhodnov I., Burko A., van den Berghe K. The Bølling–Allerød Transition in the Eastern Baltic: Environmental Responses to Climate Change // Biology. V. 12. – 2023, iss. 6. – P. 821. – Интернет-ресурс. Режим доступа: https://doi.org/10.3390/biology12060821
- Druzhinina O., Kublitskiy Y., Stančikaitė M., Nazarova L., Syrykh L., Gedminienė L., Uogintas D., Skipityte R., Arslanov Kh., , Kulkova M., Subetto D. The Late Pleistocene – Early Holocene Palaeoenvironmental Evolution in the SE Baltic Region, Kaliningrad District, Russia: a new approach based on chironomid, geochemical and isotopic data from Kamyshovoe Lake // Boreas. Vol. 49. – 2020, № 3 – P. 544–561.
- Druzhinina O., Subetto D., Stančikaite M., Vaikutiene G., Kublitsky J., Arslanov Kh. Sediment record from Kamyshovoe Lake, Kaliningrad Region: new data on history of vegetation in the late Pleistocene – early Holocene // Baltica. Vol. 28. – 2015, № 2. – P. 121–134.
- International Committee for Phytolith Taxonomy (ICPT). International code for phytolith nomenclature (ICPN) 2.0 // Annals of Botany. Vol. 124. – 2019, № 2. – P. 189–199.
- Fiłoc M., Kupryjanowicz M., Suchora M., Luoto T., Nevalainen L. Late-Weichselian (Vistulian) environmental changes in NE Poland – Evidence from Lake Suchar Wielki // Catena. – 2024, № 234. – P. 107546.
- Golyeva A. Various phytolith forms as bearers of different kinds of ecological information // Madella, M., Zurro, D. (Eds) Plants, People and Places: Recent Studies in Phytolithic Analysis. – Oxbow Books, UK, 2007. – P. 107–203.
- Mustaphi C.J.C, Pisaric M.F.J. A classification for macroscopic charcoal morphologies found in Holocene lacustrine sediments // Progress in Physical Geography. – 2014. – P. 1–21.
- Rasmussen S.O., Bigler M., Blockley S.P., Blunier T., Buchardt S.L., Clausen H.B., Cvijanovic I., Dahl- Jensen D., Johnsen S.J., Fischer H., et al. A stratigraphic framework for abrupt climatic changes during the Last Glacial period based on three synchronized Greenland ice-core records: Refining and extending the INTIMATE event stratigraphy // Quat. Sci. Rev. – 2014, № 106. – P. 14–28.
- Veski S., Amon L., Heinsalu A., Reitalu T., Saarse L., Stivrins N., Vassiljev J. Lateglacial vegetation dynamics in the eastern Baltic region between 14,500 and 11,400 cal yr BP: A complete record since the Bølling (GI-1e) to the Holocene // Quat. Sci. Rev. – 2012, № 40. – P. 39–53.
- Wachnik A. Vegetation development in the Lake Miłkowskie area, North-Eastern Poland, from the Plenivistulian to the late Holocene // Acta Palaeobot. – 2009, № 49. – P. 287–335.
