Гаплотипная структура и селекционная дифференциация пород кур по полиморфизмам гена MSTN

Автор: Дысин А.П., Дементьева Н.В.
Журнал: Молочнохозяйственный вестник @vestnik-molochnoe
Статья в выпуске: 1 (61), 2026 года.
Бесплатный доступ
В условиях растущего спроса на мясную продукцию птицы повышение эффективности селекции кур по ключевым хозяйственно-ценным признакам является актуальной задачей современной птицеводческой науки. Исследование посвящено изучению полиморфизмов и неравновесия по сцеплению гена миостатина (MSTN) в популяциях кур различной селекционной специализации. Проанализированы полиморфизмы MST_1, MST_2, MST_3 и SNP-маркеры rs317925256, rs14597164 в десяти породах кур биоресурсной коллекции ВНИИРГЖ. Установлена выраженная генетическая дифференциация между мясными, яичными и декоративными породами. Выявлено преобладание гомозиготных генотипов у мясных пород (плимутрок полосатый, пушкинская), что отражает длительную селекцию на мясную продуктивность. Полное неравновесие по сцеплению (LD = 1) между MST_1 и MST_2 и полная фиксация генотипа GG по rs14597164 обнаружены для породы китайская шелковая, что свидетельствует о продолжительной искусственной селекции. Обнаружены нарушения равновесия Харди-Вайнберга у русской белой (χ² = 8,67) и пушкинской (χ² = 9,53) пород по rs317925256, указывающие на действие направленного отбора. Итальянская куропатчатая порода показала доминирование гомозигот ТТ по rs317925256 (0,79) при высокой гетерозиготности по rs14597164 (GT = 0,63). Полученные результаты подтверждают связь полиморфизмов гена MSTN с продуктивными признаками и морфологическими характеристиками, что делает возможным использование данных маркеров в таргетированной селекции кур для повышения эффективности птицеводства.
Генотип кур, молекулярная селекция, ген MSTN, SNP-генотипирование, продуктивность птицы, равновесие Харди-Вайнберга, неравновесие по сцеплению, селекционные маркеры, генетическое разнообразие, породные различия
Короткий адрес: https://sciup.org/149150656
IDR: 149150656 | УДК: 636.5.033:575.164 | DOI: 10.52231/2225-4269_2026_1_9
Haplotype structure and selection differentiation of chicken breeds based on MSTN gene polymorphisms
With the growing demand for poultry meat products, improving the efficiency of chicken breeding for key economically valuable indicators is a relevant objective for nowadays poultry science. The research is devoted to the study of polymorphisms and linkage disequilibrium of the myostatin (MSTN) gene in chicken populations with different breeding specializations. The polymorphisms MST_1, MST_2, MST_3 and SNP markers rs317925256, rs14597164 were analyzed in ten chicken breeds from the VNIIRGH bio-resource collection. A pronounced genetic differentiation between meat, egg and ornamental breeds was determined. A predominance of homozygous genotypes in meat breeds (Plymouth Rock, Pushkin breed (Pushkinskaya)) was found, reflecting long-term selection for meat productivity. Complete linkage disequilibrium (LD = 1) between MST_1 and MST_2 and complete fixation of the GG genotype at rs14597164 were revealed for the Chinese Silk breed, indicating long-term artificial selection. Hardy-Weinberg disequilibrium was detected in the Russian White (χ² = 8.67) and Pushkin (Pushkinskaya)) (χ² = 9.53) breeds at rs317925256, pointing to the effect of directed selection. The Italian partridge breed showed a dominance of TT homozygotes at rs317925256 (0.79) with high heterozygosity at rs14597164 (GT = 0.63). The results confirm the association of MSTN gene polymorphisms with productive traits and morphological characteristics, which makes it possible to use these markers in targeted chicken selection to improve poultry farming efficiency.
Текст научной статьи Гаплотипная структура и селекционная дифференциация пород кур по полиморфизмам гена MSTN
Повышение эффективности мясного птицеводства является важным направление сельского хозяйства и требует комплексного подхода, в котором традиционные методы селекции сочетаются с современными технологиями молекулярной генетики. Применение ДНК-маркеров, в частности однонуклеотидных полиморфизмов (SNP), позволяет проводить точный отбор и значительно ускорять селекционный процесс в животноводстве [1].
Одним из ключевых регуляторов развития мышечной ткани является ген миостатина (MSTN) — член суперсемейства TGF-β. Он выступает в роли отрицательного модулятора пролиферации и дифференцировки миобластов. В птицеводстве полиморфизмы гена MSTN рассматриваются как многообещающие маркёры для генетического отбора. Исследования выявили связь между отдельными SNP и такими показателями, как прирост массы тела, убойная ценность и выживаемость у различных пород и линий кур [2, 3, 4]. Согласно современным исследованиям, ген MSTN остаётся центральным звеном в понимании молекулярных механизмов формирования мышечной массы у птиц [5]. Более того, обзорные работы подчеркивают, что MSTN является высокополиморфным геном, а его вариации могут быть использованы в качестве генетических маркеров для улучшения мясной продуктивности не только у кур, но и у других видов сельскохозяйственных животных [6, 7]. Исследования, проведённые ранее на материале генофондных пород и пушкинской популяции, подчёркивают важность изучения взаимодействия «генотип × рацион». Они также демонстрируют, что отдельные SNP в MSTN могут использоваться в качестве маркеров при отборе птиц по мясным показателям [8–11].
Тем не менее ряд вопросов остается малоизученным. Большин- ство исследований посвящено коммерческим бройлерным линиям, в то время как данные о генофондных, декоративных и гибридных породах весьма ограничены. Пока мало известно о неравновесии по сцеплению (LD) и гаплотипах внутри локуса MSTN у редких пород, закономерностях фиксации отдельных аллелей [3, 8, 11].
Цель настоящего исследования — провести комплексный анализ полиморфизма гена MSTN среди генофондных и гибридных пород кур. В рамках исследования планируется оценить частоты аллелей и генотипов по SNP rs317925256 и rs14597164, охарактеризовать локальную гаплотипную структуру, определить показатели LD и отклонения от равновесия Харди–Вайнберга, а также интерпретировать полученные результаты с точки зрения селекционной значимости и возможности реализации генетического потенциала в разных породных группах. В качестве материала для исследования были использованы образцы ДНК из биоресурсной коллекции ВНИИРГЖ, а также данные, полученные путём секвенирования фрагментов MSTN и генотипирования с применением чип-технологии Illumina Chicken 60K BeadChip.
Актуальность работы обусловлена необходимостью разработки молекулярно-генетических маркёров для селекции и сохранения генофонда.Определениепород-специфичныхпрофилейвариабельности MSTN позволит адаптировать маркерные программы (например, маркер-ассоциированную и геномную селекцию) и снизить риски потери генетического разнообразия при интенсификации селекционного отбора. Новизна исследования заключается в сочетании локального секвенирования с высокоплотным SNP-генотипированием на широком спектре генофондных и гибридных форм. Такой подход даёт возможность не только выявить ассоциированные генетические варианты, но и оценить их селекционную стабильность, а также соотнести с историей разведения конкретных пород. Теоретическая значимость работы состоит в уточнении роли гена MSTN в генетической архитектуре признаков роста у птицы. Практическая значимость заключается в разработке конкретных маркёров и рекомендаций для селекционных программ, направленных на повышение мясной продуктивности при одновременном сохранении генетического резерва.
Материалы и методы
Объектом исследования стали образцы ДНК, полученные из крови кур пород: русская белая (30 голов), пушкинская (20 голов), юрловская (20 голов), царскосельская (20 голов), плимутрок полосатый (19 голов), новопавловская (15 голов), китайская шелковая (19 голов), итальянская куропатчатая (19 голов), узбекская бойцовая х амрок (14 голов), суссекс х амрок (14 голов), брама светлая х суссекс (14 голов), царскосельская х суссекс (16 голов). Птица содержится в биоресурсной коллекции ВНИИРГЖ «Генетическая коллекция редких и исчезающих пород кур» (г. Санкт-Петербург—Пушкин). ДНК из лейкоцитов цельной крови выделяли общепринятым фенольным методом. Генотипирование животных осуществляли с помощью чипов Illumina Chicken 60KBeadChip. Биометрическая обработка данных выполнена с помощью программ Microsoft Excel, PLINK.
Результаты
В результате секвенирования гена MSTN выявлены три полиморфных сайта MST_1, MST_2 и MST_3 в группах кур пушкинская, русская белая, китайская шелковая и гибридная птица царскосельская х суссекс. Результаты попарного сравнения SNPs представлены в таблицах 1, 2, 3. Неравновесие по сцеплению равное 1 обнаружено между SNPs MST_1 и MST_2 у китайской шелковой породы (таблица 1). У той же породы не обнаружено полиморфизма по MST_3. Это говорит о наличии только двух гаплотипов по изучаемому локусу у китайской шелковой породы кур (GAC и CGC) и может объясняться историческим происхождением породы от ограниченного числа родоначальников и длительной изоляцией популяции.
Частота аллеля G у MST_2 почти по всем группам низка, кроме китайской шелковой, где этот аллель наблюдаем с частотой 0,69. Такая значительная разница в аллельных частотах может отражать различные селекционные стратегии, направленные на поддержание генетической вариабельности или на стабилизацию желательного фенотипа.
Таблица 1 – Частота встречаемости минорного аллеля и неравновесие по сцеплению между MST_1 и MST_2 маркерами (данные секвенирования по Сэнгеру)
|
Порода |
n, голов |
Частота минорного аллеля MST_1 (С/G) |
Частота минорного аллеля MST_2 (A/G) |
LD |
|
Пушкинская |
20 |
0,27 (C) |
0,06 (G) |
0 |
|
Русская белая |
16 |
0,13 (G) |
0,09 (G) |
0,02 |
|
Китайская шелковая |
10 |
0,25 (G) |
0,31 (A) |
1 |
|
Гибриды царскосельская х суссекс |
16 |
0,38 (G) |
0,03 (G) |
0,09 |
Неравновесие по сцеплению между различными парами маркеров демонстрирует неоднородность генетической структуры изучаемых популяций. Так, значение LD = 0,62 между MST_2 и MST_3 (Таблица 2) у русской белой породы указывает на сохранение определенных аллельных комбинаций, что может свидетельствовать о селекционной стабилизации благоприятных генотипов в условиях интенсивного отбора по продуктивным признакам.
Таблица 2 – Частота встречаемости минорного аллеля и неравновесие по сцеплению между MST_2 и MST_3 маркерами (данные секвенирования по Сэнгеру)
|
Порода |
n, голов |
Частота минорного аллеля MST_2 (A/G) |
Частота минорного аллеля MST_3 (C/T) |
LD |
|
Пушкинская |
20 |
0,06 (G) |
0,27 (T) |
0 |
|
Русская белая |
16 |
0,09 (G) |
0,25(C) |
0,62 |
|
Китайская шелковая |
10 |
0,31 (A) |
0 (T) |
0 |
|
Гибриды царскосельская х суссекс |
16 |
0,03 (G) |
0,31 (C) |
0,03 |
Примечательным является умеренное неравновесие по сцеплению (LD = 0,50) у гибридов царскосельская х суссекс, что отражает сохраняющиеся ассоциации аллелей, унаследованные от родительских форм. У пушкинской породы отмечено минимальное неравновесие (LD = 0,15), что свидетельствует о случайном распределении аллелей MST_1 и MST_3. Данная картина характерна для популяций, не подвергавшихся интенсивной селекции по признакам, связанным с этими конкретными полиморфными сайтами. Русская белая порода демонстрирует еще более низкое значение LD (0,11), что указывает на практически случайное распределение аллелей и отсутствие селекционного давления на комбинации именно этих маркеров. В то же время, полное отсутствие неравновесия (LD = 0) у китайской шелковой породы связано с отсутствием полиморфизма по MST_3 в данной популяции (таблица 3).
Таблица 3 – Частота встречаемости минорного аллеля и неравновесие по сцеплению между MST_1 и MST_3 маркерами (данные секвенирования по Сэнгеру)
|
Порода |
n, голов |
Частота минорного аллеля MST_1 (С/G) |
Частота минорного аллеля MST_3 (C/T) |
LD |
|
Пушкинская |
20 |
0,27 (C) |
0,27 (T) |
0,15 |
|
Русская белая |
16 |
0,13 (G) |
0,25(C) |
0,11 |
|
Китайская шелковая |
10 |
0,25 (G) |
0 (T) |
0 |
|
Гибриды царскосельская х суссекс |
16 |
0,38 (G) |
0,31 (C) |
0,50 |
Результаты генотипирования представлены в таблице 4. Анализ равновесия Харди-Вайнберга выявил значительные отклонения (χ² > 5,0) по rs317925256 у русской белой (χ² = 8,67) и пушкинской (χ² = 9,53) пород, что указывает на действие направленного отбора и дрейфа генов в этих популяциях. Указанные породы характеризуются интенсивной селекцией по мясной продуктивности, что могло привести к нарушению случайного распределения аллелей. Следует также отметить статистически значимые отклонения от равновесия у брамы светлой х суссекс (χ² = 4,54) и итальянской куропатчатой породы по rs14597164 (χ² = 2,42), что также может быть связано с селекционным давлением по фенотипическим признакам.
Таблица 4 – Частота встречаемости генотипов кур по двум полиморфным сайтам гена MSTN нек оторы х генофондны х по пуляций
|
№ n, голов |
Порода Генотипы (v) |
rs317925256 |
rs14597164 250502G/T |
χ² |
||||||
|
213016С/Т |
TT |
χ² |
||||||||
|
СС |
СT |
GG |
GT |
TT |
||||||
|
1 |
30 |
Русская белая |
0,46 |
0,23 |
0,31 |
8,67 |
0,83 |
0,17 |
0 |
0,25 |
|
2 |
20 |
Пушкинская |
0,53 |
0,12 |
0,35 |
9,53 |
0,95 |
0,05 |
0 |
0,01 |
|
3 |
20 |
Царскосельская |
0,11 |
0,31 |
0,58 |
1,39 |
0,80 |
0,20 |
0 |
0,25 |
|
4 |
19 |
Плимутрок полосатый |
0,83 |
0,17 |
0 |
0,13 |
0,42 |
0,53 |
0,05 |
0,90 |
|
5 |
15 |
Новопавловская |
0,27 |
0,46 |
0,27 |
0,07 |
0,94 |
0,06 |
0 |
0,02 |
|
6 |
19 |
Китайская шелковая |
0,11 |
0,17 |
0,72 |
2,18 |
1 |
0 |
0 |
- |
|
7 |
19 |
Итальянская куропатчатая |
0 |
0,21 |
0,79 |
0,26 |
0,32 |
0,63 |
0,05 |
2,42 |
|
8 |
14 |
Узбекская бойцовая х амрок |
0,36 |
0,36 |
0,28 |
1,11 |
0,79 |
0,21 |
0 |
0,20 |
|
9 |
14 |
Суссекс х амрок |
0,58 |
0,33 |
0,09 |
0,10 |
0,50 |
0,50 |
0 |
1,56 |
|
10 |
14 |
Брама светлая х суссекс |
0,36 |
0,21 |
0,43 |
4,54 |
0,43 |
0,57 |
0 |
2,24 |
Примечание: χ² - тест на равновесие по Харди-Вайнбергу; v – частота генотипов.
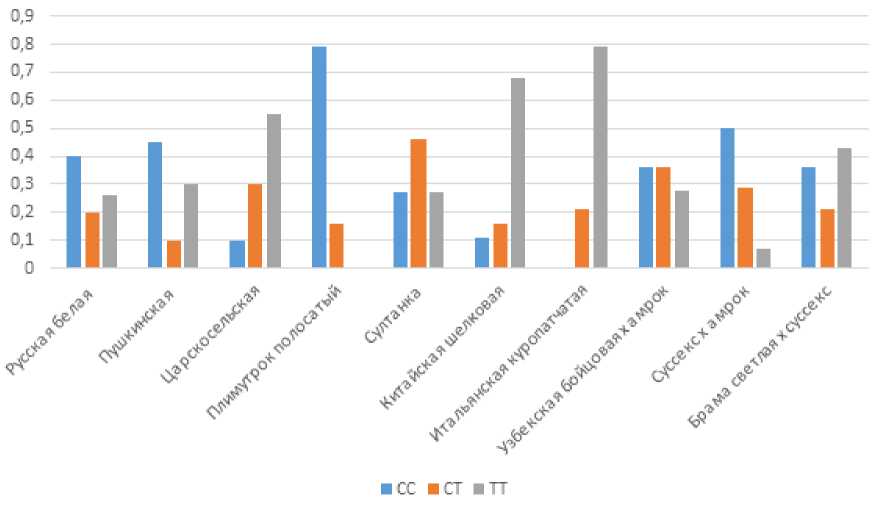
Рисунок 1 – Распределение частот генотипов изучаемых пород кур по замене rs317925256 гена MSTN
Анализ данных по замене rs317925256 (таблица 4; рисунок 1) выявляет четкие закономерности, коррелирующие с селекционной специализацией пород. Наиболее примечательным является доминирование гомозиготного генотипа СС у мясных и мясо-яичных пород. Так, у полосатого плимутрока частота генотипа СС достигает 0.83, что отражает длительное селекционное давление на мясную продуктивность в данной породной группе. Подобная тенденция наблюдается также у итальянской куропатчатой породы, где доля гомозигот ТТ составляет 0.79, что может свидетельствовать о специфических аллельных комбинациях. Особого внимания заслуживает и распределение генотипов у китайской шелковой породы, где наблюдается резкое преобладание гомозиготного генотипа ТТ (0.72) при минимальном уровне аллеля С (0.11). У новопавловской выявлена повышенная частота гетерозигот CT (0.46), что отражает отсутствие стабилизирующего отбора по локусу MSTN в декоративных линиях.
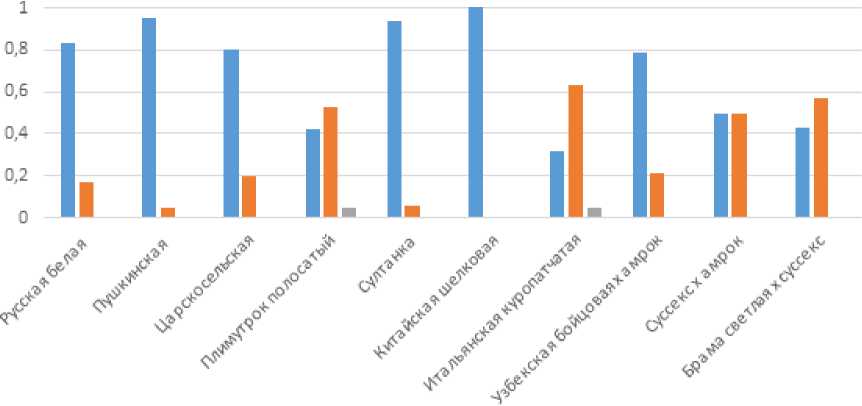
Полиморфизм rs14597164 (таблица 4; рисунок 2) демонстрирует еще более выраженную дифференциацию между породными группами. Почти во всех исследованных популяциях наблюдается доминирование гомозиготного генотипа GG, кроме итальянской куропатчатой и брамы светлой х суссекса с преобладанием GT, а также равного соотношения этих генотипов у гибридов суссекс х амрок. Особенно показательным является случай китайской шелковой породы, где генотип GG полностью фиксирован (частота 1,0), что подчеркивает отсутствие вариабельности по этому локусу и свидетельствует о древней селекционной стабилизации аллеля G. Гибридные популяции демонстрируют промежуточное распределение генотипов, что отражает эффект объединения генофондов различного происхождения и подтверждает генетическую дифференциацию исходных породных групп.
1.2
■ GG ■ GT ITT
Рисунок 2 – Распределение частот генотипов изучаемых пород кур по замене rs14597164 гена MSTN
Обсуждение
Проведенный анализ полиморфизма гена MSTN у различных пород кур выявил сложную картину генетической дифференциации, отражающую многолетнюю селекционную историю пород и специфику их использования. Наиболее показательным примером является китайская шелковая порода, демонстрирующая полное неравновесие по сцеплению (LD = 1) между маркерами MST_1 и MST_2 и одновременно отсутствие полиморфизма по MST_3. Данная генетическая структура указывает на длительную искусственную селекцию с фиксацией определенных аллельных комбинаций. Полученные результаты согласуются с данными предыдущих исследований, которые также выявляли высокую частоту полиморфных вариантов в гене миостатина у декоративных пород кур. Также, по данным Дементьевой и др. (2015), в генофондных породах наблюдаются значительные различия по частотам генотипов, особенно в участках гена миостатина, ответственных за селекционные изменения [11]. Аналогичные закономерности установлены при анализе полиморфизма в гене MSTN у кур пяти генофондных пород, где отмечено смещение в сторону преобладания определенных генотипов под действием селекционного давления.
Отдельного внимания заслуживает генетическая структура пушкинской породы, которая в нашем исследовании демонстрирует случайное распределение аллелей в большинстве локусов гена MSTN. Данный факт можно объяснить особенностями селекционной истории породы, направленной на поддержание высокой генетической вариабельности при одновременном улучшении мясных качеств. Исследования Митрофановой и др. (2019) также подтверждают динамику экстерьерных показателей у кур пушкинской породы при отборе по полиморфным вариантам в гене миостатина, что отражает сложность селекционных процессов в данной популяции [8].
Обнаруженные в нашем исследовании различия в распределении генотипов по SNP-маркерам rs317925256 и rs14597164 находят подтверждение в ряде публикаций, посвященных ассоциативному анализу полиморфизма гена MSTN с продуктивными признаками. Согласно данным Дементьевой и др. (2020), в популяциях кур мясного направления продуктивности по полиморфному варианту rs14597164 преобладает генотип ТТ (> 0,6) [12], что согласуется с нашими результатами для плимутрока полосатого, где частота генотипа TT составляет 0,05, а генотип GT доминирует (0,53). Особый интерес представляет выявленное нами нарушение равновесия Харди-Вайнберга у русской белой (χ² = 8,67) и пушкинской (χ² = 9,53) пород по rs317925256. Эти отклонения демонстрируют действие направленного отбора и дрейфа генов, что подтверждается данными исследований Ветох и др. (2023), которые показали связь генотипа GG по замене T4842G в гене миостатина с повышенными показателями живой массы у кур с разными генотипами [10]. Так, особи с генотипом GG имели значительное превосходство в живой массе.
Результаты нашего исследования по rs14597164 демонстрируют высокие частоты гомозиготного генотипа GG в большинстве исследованных популяций, что указывает на функциональную важность данного аллеля. Это согласуется с данными о связи полиморфных вариантов гена миостатина с интенсивностью роста молодняка у кур пушкинской породы, где различные аллельные варианты MSTN ассоциированы с различными показателями роста и развития [13].
Результаты анализа распределения генотипов в зависимости от селекционной специализации пород также сходятся с ранее опубликованными данными. Так, для мясных пород (плимутрок полосатый, пушкинская) характерно преобладание гомозиготных генотипов по обоим исследованным SNP-маркерам, что отражает длительную селекцию на мясную продуктивность. В свою очередь итальянская куропатчатая порода в нашем исследовании показала доминирование гомозигот ТТ по rs317925256 (0,79) при одновременном высоком уровне гетерозиготности по rs14597164 (GT = 0,63). Данная генетическая структура может отражать специфические требования к мышечной конституции. Китайская шелковая порода демонстрирует уникальную генетическую архитектуру с полной фиксацией аллеля G по rs14597164 и высокой частотой генотипа ТТ по rs317925256. Та- кие результаты согласуются с ранними исследованиями селекционной стабилизации определенных аллельных комбинаций в породах с длительной историей разведения и специфическими морфологическими особенностями [11, 12].
Заключение
Проведенное исследование комплексного анализа полиморфизма гена MSTN в популяциях кур различной селекционной специализации достигло поставленной цели по изучению генетической изменчивости в этом гене. Изученные SNP-маркеры rs317925256 и rs14597164 могут быть использованы для целенаправленной селекции кур с желаемыми характеристиками. Их включение в селекционные программы может обеспечить более точный отбор птицы и повысить эффективность отечественного птицеводства.
Перспективы дальнейших исследований включают расширение выборки для менее представленных пород, учитывание фенотипических параметров для прямого ассоциативного анализа, а также изучение влияния изученных полиморфных вариантов на экспрессию гена MSTN. Полученные данные создают основу для внедрения молекулярногенетических методов в селекционные программы домашней птицы и должны способствовать сохранению и рациональному использованию генетического разнообразия отечественных пород.
Финансирование
Работа выполнена при финансовой поддержке Министерства науки и высшего образования Российской Федерации (№НИОКТР 124020200114-7).
