Hominin-Carnivoran adaptive strategies in Western Europe during the early Pleistocene

Author: Garcia Garriga J., Martnez K., Yravedra J.
Journal: Archaeology, Ethnology & Anthropology of Eurasia @journal-aeae-en
Section: Paleoenvironment, the stone age
Article in issue: 2 т.44, 2016.
Free access
Short address: https://sciup.org/145145260
IDR: 145145260 | DOI: 10.17746/1563-0110.2016.44.2.019-029
Text of the article Hominin-Carnivoran adaptive strategies in Western Europe during the early Pleistocene
The first peopling of Europe is still the focus of heated debate, with a number of researchers considering this issue from different perspectives. In some cases, the interest has been focused on its chronology (Muttoni, Scardia, Kent, 2013; Garcia et al., 2014); although possible routes (Rook, Martínez-Navarro, Howell, 2004), the variables conditioning the expansion towards Europe and Asia (Turner, 1992; Arribas,
Palmqvist, 1999), the importance of the climate and the environmental conditions (Rodríguez-Gómez et al., 2014), and the continuous or discontinuous nature of these occupations have been also discussed (Garcia, Martínez, Carbonell, 2011; Bermúdez de Castro et al., 2013). It has been suggested that during the Early Pleistocene, hominin settlement was sporadic, resulting from several migratory events that alternated with episodes when the continent was under-populated, or even deserted (MacDonald et al., 2012). The limited number of sites and the paucity of hominin fossils between 1.4 and 0.7 Myr, together with the assumed inability of hominins to colonize the lands north of the 50º N latitude—although disputed by later findings of anthropic activity at Happisburgh 3, in England (Parfitt et al., 2010) and Untermassfeld, in Germany (Landeck, 2010; Garcia et al., 2013; Landeck, Garcia Garriga, 2015; Landeck, Garcia Garriga, 2016)—are some of the arguments supporting this hypothesis. Nevertheless, the palaeoecological stability recorded in southern Europe during the Early and Middle Pleistocene, as evidenced in the Sierra de Atapuerca and the archaeological sequences at Barranco León D and Fuente Nueva 3 (Orce), Vallparadís (Barcelona), Sima del Elefante and Gran Dolina TD6 (Atapuerca), points to a continuous hominin occupation.
The archaeological record of the European Early Pleistocene is increasingly better represented, thus providing the needed documental basis to interpret the causes, evolution and fundaments of the first peopling of Europe. The pre-Jaramillo lithic industry and/or fossil remains found at Barranco León D and Fuente Nueva 3, together with those from Sima del Elefante, Pirro Nord and Kozarnika cave, prove the presence of hominin groups by 1.4–1.1 Myr. Hominin evidence roughly corresponding to the Jaramillo subchron has been documented at Untermasffeld, Ca’Belvedere di Monte Poggiolo, Pont de Lavaud, Lunery-Rosières, and Le Vallonnet. After the Jaramillo episode, between 0.98 and 0.78 Myr, the extent and quality of the record appears significantly enlarged, contributing new data for this long period with sites like Vallparadís, Gran Dolina TD6, Pakefield and Happisburgh 3, Soleilhac, and Korolevo (See Figure and Table ). All these sites support a continuous hominin occupation, probably from 1.4 Myr to the Matuyama-Brunhes boundary (Garcia, Martínez, Carbonell, 2011; Garcia et al., 2014). This hypothesis contradicts the thesis supported by Muttoni et al. (Muttoni, Scardia, Kent, 2013), who argue that the first European settlement should be chronologically constrained to 0.85 – 0.78 Myr.
According to zooarchaeological and taphonomic data, the hominins peopling the mid/low latitudes had access to a wide range of prey, which may have facilitated their adaptation to European ecosystems (Martínez et al., 2010; Rodríguez et al., 2011; Saladié et al., 2011, 2014; Huguet et al., 2013; Landeck, Garcia Garriga, 2016). Despite their primary access to herbivore carcasses, before they dispersed out of Africa about 1.8 Myr, being unquestionable and well-known, there is no consensus amongst researchers about hominins’ place in the trophic chain regarding large carnivorans (i.e. Pachycrocuta brevirostris , Panthera gombaszoegensis , and Homotherium latidens ). Taking into account the taphonomic evidence provided for the ‘hunting versus scavenging’ debate, we discuss here the implications of hominin-carnivoran competition, and their possible consequences for the continuity or discontinuity of the first hominin settlement in the mid/ low latitudes of Europe.
Materials and methods
This paper addresses the study of the hominin peopling of Europe during the Early Pleistocene, considering the zooarchaeological and taphonomic data recovered from the most relevant sites in the south-west of the continent (See Figure and Table ). Our aim is to confirm whether the subsistence strategies regarding animal foodresources adopted by the first European hominins were dependable enough to justify a continuous population. On the other hand, owing to our interest in analyzing hominin activity on the fauna, we will focus exclusively on the sites with zooarchaeological and taphonomic data, as this information allows evaluation of the degree of intervention of hominins on bone accumulations (see Table ).
The Early Pleistocene in the Iberian Peninsula
Fuente Nueva 3 and Barranco León D (Orce), Sima del Elefante and Gran Dolina TD6 (Atapuerca), as well as Vallparadís, may be classified as Type C sites according to Isaac (1983), because they present a lithic industry associated with several herbivore individuals. However, this association does not necessarily imply a direct or functional relationship between the different remains: numerous sites exemplify accidental associations of fauna and lithic industries that originated in independent events. Thus, in order to
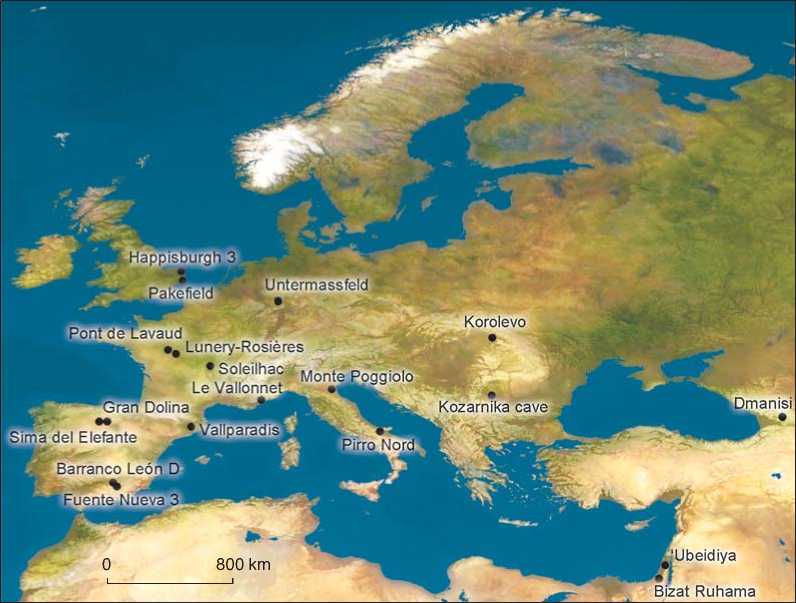
Geographical situation of the Eurasian and circum-Mediterranean archaeological sites mentioned in the text.
relate the faunal remains found in a site to a hominin agent, it is necessary to study the sedimentary and taphonomic processes responsible for site-formation. Hence, the identification of cut-marks and percussionmarks, and their representation and anatomical distribution, are unmistakable arguments in favor of hominin intervention on a carcass. The analysis of these alterations, together with other patterns such as fracture mechanisms or mortality profiles, helps identify in which order hominins had access to a carcass, and the kind of intervention performed.
The Guádix-Baza basin
The Guádix-Baza basin (southern Spain) includes several paleontological and archaeological sites of different chronologies. The sites presenting a clear association of lithic artifacts and faunal remains are Fuente Nueva 3 and Barranco León D, which are dated at 1.4–1.2 Myr (Toro-Moyano et al., 2009; Espigares et al., 2013). At Barranco León D, some herbivore remains yielded cut-marks and spiral or helical fractures, as well as impact-points, flake scars and bone flakes. For example, a rib-fragment of Hippopotamus antiquus shows a cut-mark which is related to evisceration processes (Toro-Moyano et al., 2013). In the upper archaeological level of Fuente Nueva 3, an incomplete elephant-carcass was found associated with 34 coprolites of Pachycrocuta and 17 lithic tools (Espigares et al., 2013). This record suggests that hominins and carnivorans may have gotten access to this animal in turns. However, no direct evidence relates the proboscidea with either hominin or carnivoran consumption. The elephant bones did not yield any cut-marks, percussion-marks or fractures. However, their association with the lithic tools may imply hominin intervention on the carcass.
The upper and lower archaeological levels at Fuente Nueva 3 present two rather similar assemblages. The upper level denotes a marginal occupation by hominins, whereas the lower levels yielded Mode 1 tools and some large mammal (between 340 and 907 kg) tibiae and radii with defleshing and disarticulating cut-marks (Ibid.). The reduced number of remains with anthropic traces prevents the identification of hunting or scavenging strategies in the lower levels at Fuente Nueva 3; although some of the cut-marks on the pelvis, ribs and femur may suggest an early access to the carcasses by hominins before the intervention
Characteristics of the faunal record in the main archaeological sites of Western Europe in the Early Pleistocene
|
tn 0) о с Ф т |
CD со со g со 5 ^ " см о <м 2° со см га со со m 2 g CD ” CD СМ с СМ °Ф § О О CD О ” -Г СО _Г с £ см . ” 5 5 - ф ” ””Б * 5 2 о о "55 ® ф Ф 2 см g с ™ ф 2 g 2 й ® го ^ о 8 -1 jo ф g СП бел о-g .ТОГО У СУ с О е о Е Е ГО Го т ГО ГО CXJ CXJ О = О LL ГО CXJ F F О СО ТО ш > со О |
|
О § g^ |
™ С о 2 ф .с ф с § ф ф
о о о о со о о _ Н ГО С £ с ГО го CD с 9 ф > Р ф Р о Е о Е > о го > го у Г- ф с Ф г- ь— с о го го -с с>.Е го с о о о -с о -с
|
|
о н о Е СО С о о ГО |
” S Го 1 е I ■ 2 с ф > 2 Е ф 2 о 2 £ го Е -с £ Е £ го Е £ £ го £ с о го го го го с о го ГОССС < (У) > z О < о z < < |
|
tn го "Е го О |
+ + + + + + + + + го + |
|
tn го Е ф m |
5^ + + -о ш + + + 2 о + + + + 00 § Н О СО О" < — |
|
« с 2 ел со И о а |
го + + го ш + i^ S + 2 ^ + । । + со У Н СО О *М ^ |
|
с О X го с 'го |
Р "О СО -О с Ф с -о р го го го го со О § ■О 2 Р Р „ 0 2 8-0 о- ф ф ф о- ф ф Р ош О О Ош QQ £ < О О- ш |
|
с го О |
ГО ф Го Q) с - > -С - > ф - Го - ф ' ГО О О О О |
|
о ^ О О g. б |
CN CN |^ V V cn о о о ” о со 4 4 ^ 6 6 О 05 О ^ ^ о |
|
tn ф О |
00 Q £ со ю > со ГО С Q н Q ф -О S ш н Ш ел ® g 2 CQ (Z) О О > _| 2 СЛ о |
of carnivorans (Ibid.). As well as defleshing and disarticulating cut-marks, percussion-marks are observed on the diaphyses of some radii, femora and tibiae, indicating marrow extraction. Hence, an intensive exploitation of the carcasses is suggested: first the flesh and later the marrow were equally consumed. At the lower levels, carnivoran action is also reflected in the pits and scores recorded on some of the bones. Owing to their low numbers, low carnivoran activity on the fauna can be advanced for the lower levels at Fuente Nueva 3, as opposed to the upper levels which yielded a record of tooth-marks and Pachycrocuta coprolites.
The Sierra de Atapuerca
In the Sierra de Atapuerca (Burgos, northern Spain) hominin evidence from the Early Pleistocene was recovered at Sima del Elefante and Gran Dolina (Bermúdez de Castro et al., 1997; Carbonell et al., 2008). Together with hominin remains, Mode 1 lithic tools were associated with fauna (Carbonell et al., 1995, 2008). The cut-marks identified indicate that the bones were processed by hominins, including some of the hominin remains (Rodríguez et al., 2011; Saladié et al., 2011, 2014; Huguet et al., 2013, in press). The different levels from Sima del Elefante were subject to different taphonomic processes (Rodríguez et al., 2011; Huguet et al., 2013, in press). In level TE9, the cut-marks on several deer-, bison- and turtle-bones correspond to skinning, dismembering and defleshing processes (Ibid.). The presence of cut-marks on axial and appendicular bones suggests a primary access to the carcass by hominins. However, the reduced number of remains identified hampers any conclusion regarding the reliability or peculiarities of this intervention. Furthermore, carnivoran-altered bones were recovered from all levels. Even though they are not numerous, their presence indicates some degree of competition between hominins and carnivorans for access to the carcasses. However, with the reduced record available, it is not possible to identify the agent responsible for accumulating the bone assemblages found in Sima del Elefante; although the presence of some cut-marks typical of defleshing may suggest that at least some of the animals were introduced by hominins.
In Gran Dolina, hominin activities also vary according to the levels. In TDW4, the bones were naturally assembled as consequence of collapsed boulders and bear-hibernation. However, marks of evisceration and defleshing have been registered on meat-bearing bones that can be considered as typical traces of primary flesh-exploitation by hominins. Similarly to the rest of the sites considered, there is also evidence of carnivoran activity representing 9.2 % of the sample (Huguet et al., 2013). Regarding Gran Dolina TD6, both skeletal profiles and bone alteration patterns indicate that, despite hominin-carnivoran competition, hominins had access to flesh and were the main agents responsible for the transportation of the different animals to the site (Díez et al., 1999; Saladié et al., 2011, 2014; Huguet et al., 2013; Rodríguez-Gómez et al., in press). Both the amount of cut-marked bones (13 %) and the overlapping of toothmarks on cut-marks (Huguet et al., 2013) support this interpretation. The different traces indicate unrestricted access to the carcasses and the performance of several activities such as dismembering, skinning, evisceration and defleshing, which would eventually deplete the carcasses. Díez et al. (1999) indicate the differential transportation of prey according to their size, whereby small- and medium-sized animals were introduced in complete condition to the sites. Nevertheless, Saladié et al. (2011) add that some large-sized prey were also complete on arrival, a fact which may imply cooperation and sharing of resources by the group.
Carnivoran activity has also been documented in Gran Dolina TD6. Some overlapping of tooth-marks on cut-marks indicates that carnivorans scavenged the remains left by hominins (Díez et al., 1999; Saladié et al., 2011; Huguet et al., 2013), and that they played a significant role in the damage of the ungulate carcasses in TD6-2 after hominins had extracted a large amount of meat (Saladié et al., 2014). Together with the exploitation of these ungulates, numerous remains of Homo antecessor bearing cut-marks have been documented in TD6, constituting the first evidence of cannibalism known in human evolution. This behavior is interpreted as regular cannibalism with nutritional purposes (Carbonell et al., 2010). Nevertheless, the record found in Gran Dolina TD6 indicates that hominins were able to gain primary access to carcasses. Further studies have confirmed that the hominin population’s age profiles from this level are similar to those seen in cannibalism practices resulting from intergroup aggression among chimpanzees (Saladié et al., 2012). This cannibalism may have been an adaptive strategy to alimentary stress-situations, a practice aiming at preventing carnivorans from scavenging hominin bodies once the site was abandoned, or may have resulted from intergroup competition. Whatever the case, it is significant that a large number of ungulates were hunted contemporaneously with the cannibalistic consumption of hominin remains; which may imply that this practice did not just respond to an alimentary need despite the economization of resources suggested by the consumption of hominin bodies.
Vallparadís
The richest archaeological unit of Vallparadís (Barcelona, north-eastern Spain) is unit EVT7 (archaeological levels 10 and 10c). It is 3 m deep and was completely excavated, covering over 780 m2. This unit has been chronologically dated by the combination of biostratigraphic analyses of micro-and macromammals, palaeomagnetism, ESR, and OSL radiometric dates. It presents a negative polarity corresponding to an age between the Jaramillo event and the Matuyama-Brunhes boundary. The association with Mimomys savini and Iberomys huescarensis rodents further situates Vallparadís in the Allophayomis chalinei biozone (Martínez et al., 2010). A comparison of the mandibular first molars of individual adult specimens of Mimomys savini recovered from unit EVT7 with specimens from the stratigraphic sequence at Gran Dolina, Fuente Nueva 3, and Barranco León D has provided a chronology of around 0.98–0.95 Myr (MIS 27) (Martínez et al., 2014; Garcia, Martínez, 2015). Additionally, ESR and OSL analyses on equid teeth and quartz grains indicate mean dates of 0.83 ± 0.13 Myr and 0.849 ± 0.48 Myr for archaeological level 10, and an age of 0.858 ± 0.87 Myr for unit EVT8 (Martínez et al., 2010; Duval et al., 2011, 2015).
Evidence in favor of the presence of hominin activity at the site and primary position of part of the archaeological record is discussed in Garcia et al. (2012). A rich Mode 1 bipolar-on-anvil lithic assemblage was found at EVT7 (Garcia, Martínez, Carbonell, 2013), in association with fossil herbivoreremains with cut-marks and anthropic fractures. The vertical distribution of the archaeological record at this unit yielded three differentiated assemblages of lithic and faunal remains, suggesting the occurrence of several sedimentation events. Together with the identification of four lithic refittings, this sequence may indicate the primary position of part of the record. The macrofaunal assemblage collected from unit EVT7 may be described as Epi-Villafranchian. The taphonomic analysis of the bone remains indicates that the scavenger Pachycrocuta brevirostris was the main agent responsible for the assemblage: many of the bones present gnawing-marks, cylinders are abundant (Madurell-Malapeira et al., 2012), and some regurgitated bone-fragments and coprolites have also been identified (Martínez et al., 2010; Garcia et al., 2012).
Hominin intervention on herbivore carcasses has been evidenced on several bones of very large animals such as Rhinocerotidae and Hippopotamidae, as well as on the remains of medium-sized herbivores such as Equidae and Cervidae. Twelve elements from the upper limbs, axial bones, and lower appendicular bones represent dismembering, defleshing, and skinning actions; and five elements show fractures of anthropic origin (Martínez et al., 2010; Garcia et al., 2012). Half of the bones with cut-marks also present tooth-marks, which indicates that both hominin and large predators gained access to the same prey. Marks produced by both agents on the same bones could imply competition between hominins and large predators for the carcasses; or, alternatively, facilitation of the resources, as the scavenger may have consumed the remains abandoned by the earlier predator. No overlapping marks have been found so far to define the access sequence to the flesh by both agents. The low frequency of anthropic marks, and the notorious carnivoran activity, suggests a low participation of hominins in a site mainly inhabited by Pachycrocuta brevirostris .
Other areas: Italy and France
There are some other sites from the late Early Pleistocene which may provide data on the first peopling of Europe. Soleilhac (France) yielded an important faunal collection of more than 3000 remains from 29 individuals associated with lithic industry (Bonifay, Bonifay, 1981). Although hominins may have been responsible for the accumulation of most of the bone-assemblage, these remains are poorly preserved, hampering their identification. Only the identification of deer-bones seems to be conclusive (Fosse, 1994). In Le Vallonnet (France), remains from the early Middle Pleistocene were also recovered. Despite doubts about its associated lithic industry, the abundant fauna of the site indicates a low anthropic impact that may have focused only on cervids (Echassoux, 2009).
Turning to Italian sites, ancient evidence of occupation has been found in Pirro Nord; but no zooarchaeological data are currently available. Other sites such as Ca’Belvedere di Monte Poggiolo have reported some contextual issues related to the association of fauna and lithic artifacts (Villa, Lenoir, 2009). Although in these sites hominin activity is proven by the presence of lithic artifacts and cut-marks on some bones, other sources of evidence also report important participation by carnivorans. In Le Vallonnet (Echassoux, 2009) and Pirro Nord (Arzarello et al., 2012) some remains of Pachycrocuta are documented. Their presence suggests that hominins and carnivorans may have competed for the access to the carcasses or, alternatively, that hominins were exposed to the limitations imposed by carnivorans.
Discussion
Available zooarchaeological and taphonomic evidence for the Early Pleistocene in the Iberian Peninsula indicates that hominins had access to a variety of meat resources, from small animals such as birds and reptiles to large mammals like hippopotamuses and ungulates of varying sizes—i.e. cervids and bovids (Martínez et al., 2010; Rodríguez et al., 2011; Saladié et al., 2011, 2014; Huguet et al., 2013, in press) (see Figure and Table ). Evidence of carnivoran activity is frequently found contemporaneously with hominin intervention at the Sima del Elefante, Gran Dolina, Orce, and Vallparadís sites. Owing to the incompatibility of hominins and carnivorans coexisting in the same site, the presence of both agents indicates a typical situation of stress and competition for resources and the occupation of the same spaces throughout the Early Pleistocene. Because of this relationship between hominins and carnivorans, some authors have envisaged the first peopling of Europe on the basis of the presenceabsence of carnivorans. According to this view, hominins from Africa would emigrate by scavenging the carcasses abandoned by carnivorans (Turner, 1992; Arribas, Palmqvist, 1999). In other cases, the hominin expansion to Eurasia was favored by the extinction of the Pachycrocuta species, which would reduce trophic competence and allow hominins to gain access to a larger number of carcasses (Turner, 1992). On the other hand, some authors consider that carnivoran trophic pressure did not have any influence in hominin behavior and adaptability to the different areas that were gradually colonized (Echassoux, 2009).
Carnivoran presence in Europe was constant throughout the Pleistocene, although their representation over time was uneven. At the beginning of the Early Pleistocene, the ratio of carnivorans to herbivores was quite high, with a large number of hyperspecialized carnivorans. This carnivoran representation was increased by the arrival of new species such as Lycaon, Pachycrocuta brevirostris, Canis etruscus, or Ursus etruscus, which were better adapted to open dry landscapes (Croitor, Brugal, 2010). From the Early to Middle Pleistocene transition, solitary hypercarnivorans such as Acynonix, Megantereon, Dinofelix, Homotherium or Lynx issidorensis were extinguished, giving place to smaller carnivorans with more generalized and gregarious behavior (Ibid.). In Gran Dolina TD6, both the faunal representation and the percentage of cut-marked bones are important, affecting several taxa and all bone parts. The cut-marks identified indicate butchering, evisceration, skinning, and disarticulation operations. Additionally, ageprofiles reveal the dominance of young animals. This evidence suggests a prominent anthropic participation in carcass accumulation, further confirmed by overlapping of tooth-marks on hominin-inflicted traces, indicating secondary carnivoran intervention on the carcasses (Saladié et al., 2011, 2014; Huguet et al., 2013). It may thus be concluded that at this level, hunting strategies could have been used regularly, and prey were transported to the site to be shared with the rest of the group.
Furthermore, it should be noted that in other sites, meat resources were obtained in alluvial and fluvial environments that were hostile to hominins, given the high level of competition with carnivorans; such is the case with Fuente Nueva 3, Barranco León D, and Vallparadís. In these cases, the early or primary hominin access to meat resources can be interpreted in the presence of cut-marks on the meat-bearing bones of medium-sized animals such as deer. Experimental studies show that these traces are not frequent on the more nutritious elements after carnivoran intervention (Gidna, Yaravedra, Domínguez-Rodrigo, 2012). This primary hominin access to the carcasses in the European Early Pleistocene, exemplified in Gran Dolina TD6, is related to high trophic pressure in several areas. Thus, the cut-marks identified by Espigares et al. (2013) on the ribs, the pelvis, and the diaphysis of the femur in the lower levels at Fuente Nueva 3, as well as the cut-marked bones from the upper limbs, axial, and lower appendicular parts studied by Martínez et al. (2010) in Vallparadís, are incompatible with scavenging behavior by hominins, providing evidence for their early access to the carcasses.
As regards the order in which hominins and/or carnivorans gained access to the carcasses in Vallparadís and Fuente Nueva 3, it is not possible to specify the sequence, because of the absence of overlapping marks. Differences in taphonomic data at the sites may be related to their location, as well as their specific function. So, both the sites at Orce and Vallparadís are open-air occupations near a water source; while Gran Dolina TD6 is a cave site where intensive campsite- like occupations have been documented (Saladié et al., 2011, 2014; Rodríguez-Gómez et al., in press). At Gran Dolina TD6 there is a higher percentage of cut-marked bones and a lesser degree of carnivoran intervention, while fossil bones are more affected by large predators at the fluvio-lacustrine sites of Orce and Vallparadís— together with some sporadic evidence of anthropic intervention, an activity which may also have partially destroyed anthropic evidence.
Conclusions
The archaeological sequence drawn from the sites in Orce, Atapuerca, and Vallparadís covers a period extending from 1.4–1.2 Myr to the Matuyama-Brunhes boundary (0.78 Myr). Their long chronology suggests that hominin occupation was continuous in the Iberian Peninsula (Garcia et al., 2011, 2014). Taphonomic and zooarchaeological analyses indicate that in all of these Iberian sites—as in the Eurasian sites—the same kind of basic adaptive strategy was adopted, characterized by cohesive groups capable of obtaining meat resources regularly (Martínez et al., 2010; Saladié et al., 2011, 2014; Huguet et al., 2013, in press). Furthermore, all of them present lithic assemblages with comparable characteristics, associated with a Mode 1 technocomplex of some variability (Garcia et al., 2013). Such adaptive and technological features may have offered these populations the possibility of adapting to the different ecosystems and climatic fluctuations recorded throughout this period.
The faunal evidence recovered from the European sites shows a high number and a large variability of ungulate taxa, thus providing hominins with an abundance of faunal resources. On the other hand, the ratio of carnivorans to herbivores decreases towards the end of the Early Pleistocene (Coritor, Brugal, 2010). Both situations may have favored hominin dispersion throughout the European continent during the Early Pleistocene. The evidence from the Iberian sites discussed, as well as that from the rest of Europe, demonstrates that the consumption of animal flesh was an early strategy. Eurasian sites such as Gran Dolina TD6 (Díez et al., 1999; Saladié et al., 2011, 2014; Huguet et al., 2013; Rodríguez-Gómez et al., in press), Fuente Nueva 3 (Espigares et al., 2013; Huguet et al., 2013), and some levels of ‘Ubeidiya (Belmaker, 2006) and Bizat Ruhama (Zaidner, Yeshurun, Mallol, 2010) have reported clear evidence of hominin activity. In Gran Dolina TD6 this evidence may prove primary access to animals (including hominin specimens)
weighing between 10 and 400 kg for their flesh, probably by hunting, or, alternatively, by scavenging large animals such as elephants or hippopotamuses (Díez et al., 1999; Saladié et al., 2011, 2014; Huguet et al., 2013).
Other sites, such as Sima del Elefante and Barranco León D (Huguet et al., 2013), Vallparadís (Martínez et al., 2010; Garcia et al., 2011, 2012, 2014), Untermassfeld (Landeck, Garcia Garriga, 2016), and Dmanisi (Tappen et al., 2007), also present evidence of meat exploitation by hominins; but significant action by carnivorans cannot be disregarded. In most cases, hyaenids (Ibid.; Madurell-Malapeira et al., 2012) and felids (Arribas, Palmqvist, 1999) would be mainly responsible for the faunal assemblages. This implies that hominins and carnivorans were forced to compete for the same natural resources and, in order to expand throughout Europe and Asia, hominins had to face serious challenges such as trophic pressure and adaptation to new environments.
The consumption of large animals such as hippopotamuses in sites like Vallparadís, ‘Ubeidiya, Untermassfeld, or at Orce may have responded to a sporadic exploitation pattern. The presence of these large-sized carcasses in their typical environment may suggest their natural death or, alternatively, their hunting by large felids, to be subsequently exploited by hominins. Even if they responded to a secondary consumption, it also implies direct competition with large hyaenids for the exploitation of large carcasses, giving hominins a more active role as scavengers than that described by Turner (1992). The greater climatic stability in the southern refugia such as the Iberian Peninsula, and the evidence of anthropic modifications present on the prime meat-bearing parts of large herbivore carcasses, might indicate that these early groups were able to establish continuous settlement in the Iberian Peninsula and to expand to northern Europe in warmer episodes.
