К вопросу о древнейших следах появления человека на юге Восточной Европы (Россия)
")
Автор: Саблин М.В., Гиря Е.Ю.
Журнал: Археология, этнография и антропология Евразии @journal-aeae-ru
Рубрика: Палеоэкология. Каменный век
Статья в выпуске: 2 (42), 2010 года.
Бесплатный доступ
Фрагмент плюсневой кости древнего верблюда Paracamelus alutensis со следами рубки и резания обнаружен в нижних горизонтах хапровского аллювия. Согласно результатам анализа общей морфологии, следы оставлены одним орудием с достаточно массивным и острым лезвием. Это мог быть либо чоппер, либо массивный отщеп. Результаты исследования не оставляют сомнений в том, что следы рубки и резания появились в процессе разделки свежей туши древнего верблюда Paracamelus alutensis - типичного представителя хапровской фауны. Стратиграфический диапазон распространения хапровского фаунистического комплекса определяется финалом среднего виллафранка в интервале 2,1-1,97 млн лет. Контекст обнаружения кости древнего верблюда Paracamelus alutensis, степень ее сохранности и выразительность следов работы каменным орудием, оставленных на поверхности, могут свидетельствовать о достаточно раннем появлении человека в Евразии.
Следы рубки, трасология, эксперимент, хапровская фауна, стратиграфия, средний виллафранк
Короткий адрес: https://sciup.org/14522792
IDR: 14522792 | УДК: 551.782.23
Текст научной статьи К вопросу о древнейших следах появления человека на юге Восточной Европы (Россия)
Первые бесспорные следы пребывания человека на Кавказе в олдованскую эпоху были найдены более 20 л.н. [Dzaparidze et al., 1989]. Однако результаты работ только последних семи лет позволили значительно расширить географию и хронологию археологических источников наших знаний о человеке этой эпохи в Кавказском регионе и на сопредельных территориях.
В 2002 г. Кубанской палеолитической экспедицией Института истории материальной культуры РАН был открыт комплекс нижнепалеолитических стоянок Бо- гатыри – Родники на Тамани [Кулаков, Щелинский, 2004; Bosinski et al., 2003]. С 2003 г. в Дагестане ведут систематические исследования экспедиция Института археологии и этнографии СО РАН под руководством акад. А.П. Деревянко и Северокавказская палеолитическая экспедиция Института археологии РАН, возглавляемая чл.-кор. РАН Х.А. Амирхановым. В 2003– 2005 гг. там были выявлены десятки стратифицированных памятников олдованской и раннеашельской эпох [Амирханов, 2007; Деревянко и др., 2009].
Следствием этих открытий стало резкое изменение объема и качества археологических источников. Появилась новая парадигма, с которой связано начало нового этапа в истории отечественного палеолитоведения. За короткий период изменились не только общие представления о хронологии и географии древнейшего палеолита. Внутри этой эпохи теперь уверенно выделяются вполне обособленные типы индустрий, различные как по технологии расщепления, так и по
типам изделий [Деревянко, 2009]. По мнению специалистов, «примитивизм» древнейших орудий не означает отсутствие достаточно четко выраженных типов [Амирханов, 2006; Таймазов, 2009; Щелинский, Кулаков, 2009]. Более того, многие вопросы генезиса древнейших культур вполне успешно решаются «в плоскости сравнительно-исторической типологии» [Амирханов, 2007, 2009; Любин, Беляева, 2006].
Сегодня древнейшими на Кавказе и во всей Евразии признаны находки из Дманиси – им ок. 1,8 млн лет. Недавний пересмотр палеонтологических коллекций ЗИН РАН позволяет констатировать еще более раннее появление человека на евразийском континенте.
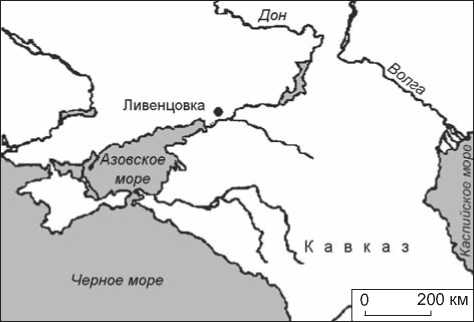
Рис. 1. Местонахождение Ливенцовского карьера.
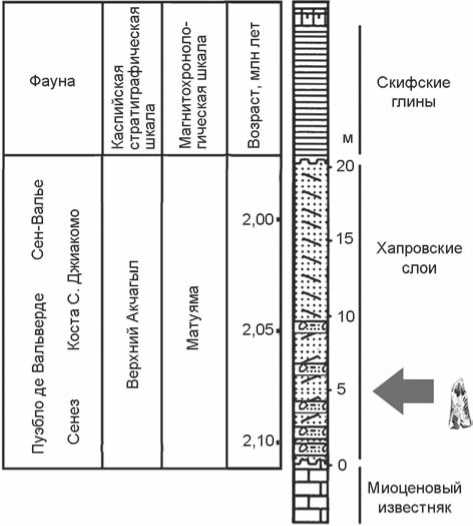
Рис. 2. Схематический разрез Ливенцовки.
Исследуемый фрагмент кости № 35676 из коллекции ЗИН РАН был найден вместе с другими фаунистическими о статками в хапровском аллювии Н.К. Верещагиным при посещении им в мае 1954 г. Ливенцовского карьера, но не был тогда опознан как артефакт. Карьер находился на западной окраине г. Ростова-на-Дону, в настоящее время рекультивирован. Местонахождение Ливенцовка (координаты: 47° 13´ с.ш., 39° 34´ в.д.; рис. 1) признано парастратотипом хапровского фаунистического комплекса, выделенного В.И. Громовым [1939]. Хапровские слои являются аллювием террасы Дона; в них большинство исследователей выделяет две пачки отложений суммарной мощностью до 20 м. Нижняя пачка мощностью 7–9 м содержит грубообломочный материал и кости крупных млекопитающих [Байгушева, 1971; Александрова, 1976]. Именно в нижних горизонтах аллювия, как следует из записей Н.К. Верещагина, были обнаружены фаунистические остатки (рис. 2).
Всего же в хапровских слоях, относящихся к русловой фации палео-Дона, разными исследователями было собрано более 3 тыс. определимых костных фрагментов сходной степени фоссилизации и принадлежащих 33 видам крупных млекопитающих [Титов, 1999, 2008]. Преобладают изолированные кости животных, но известны и целые черепа и части скелета в естественном сочленении. Ископаемый материал коричневого или светло-серого цвета, сохранность основной массы находок одинакова. Формирование ориктоценоза происходило в течение одного седиментационного цикла палео-Дона, что позволяет считать хапровский фаунистический комплекс крупных млекопитающих хронологически единым. Данная ассоциация существовала в условиях жаркого сухого климата саванны: в сборах палеонтологов доминируют остатки лошади Equus livenzovensis (24 %), южного слона Archidiskodon meridionalis (23 %), верблюда Paracamelus alutensis (13 %), встречаются кости страуса Struthio , эласмотерия Elasmotherum , древнего жирафа Palaeotragus [Титов, 1999, 2008; Bajgusheva, Titov, Tesakov, 2001; Bajgusheva, Titov, 2004]. О сухом и жарком климате в период формирования кос-тено сных горизонтов хапровского аллювия свидетельствуют минералогические характеристики его компонентов, а также присутствие в русловой фации палео-Дона раковин теплолюбивого пресноводного моллюска Bogatschevia tamanensis [Разрез…, 1976]. Время формирования аллювия исследователи, изучающие ископаемых моллюсков, относят к верхнему акчагылу [Rudyuk, 2003]. Это в совокупности с обратной намагниченностью всей 20-метровой толщи хап-ровских слоев [Tesakov et al., 2007] свидетельствует о доолдувейском возрасте отложений.
Ассоциация грызунов Borsodia newtoni , Borsodia arankoides , Mimomys reidi , Clethrionomys kretzoii ,
Mimomys pliocaenicus , о статки которых собраны в верхних 15 метрах хапровского аллювия, по степени эволюционного развития полевок отнесена к концу зоны MN17 и помещается в магнитохронологической шкале между верхней границей эпизода Реюньон и нижней границей эпизода Олдувей [Тесаков, 2004]. Присутствие в составе хапровской фауны таких видов крупных млекопитающих, как Canis cf. senezensis , Nyctereutes megamastoides , Ursus cf. etruscus , Sus strozzii , Libralces gallicus , Leptobos cf. etruscus позволяет синхронизировать ее с европейскими фаунами из местонахождений Пуэбло де Вальверде, Сен-Валье, Сенез, Коста С. Джиакомо. Согласно новейшим исследованиям, все эти фауны располагаются в магнитохронологической шкале между верхней границей эпизода Реюньон и нижней границей эпизода Олдувей [Сотникова, Бай-гушева, Титов, 2002; Torre et al., 1992; Roger et al., 2000; Channell, Labs, Raymo, 2003; Guerin et al., 2004; Sinusia et al., 2004]. Таким образом, стратиграфический диапазон распространения хапровского фаунистического комплекса определяется финалом среднего виллафранка в интервале 2,1–1,97 млн л.н.
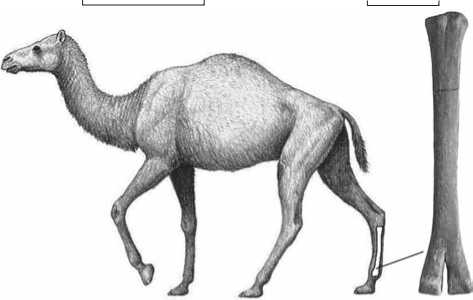
Исследуемый артефакт (длина 106,5 мм, ширина 49,0 мм) представляет собой дистальный фрагмент задней правой метаподии (metatarsus) древнего верблюда Paracamelus alutensis – типичного представителя восточно-европейских фаун конца плиоцена (рис. 3). Всего в хапровском аллювии было найдено более 200 костей этого животного. Морфологический анализ данного материала [Титов, 2005; Titov, 2003] показал, что Paracamelus alutensi s, обладавший стройной шеей и грацильными конечностями, пропорциями тела напоминал современную южно-американскую ламу, а по высоте в холке не уступал рецентным одногорбому и двугорбому верблюдам. Внутри костномозгового канала metatarsus сохранился характерный для хапровских слоев белый уплотненный кварцевый песок, в котором, вероятно, и была погребена кость. Это исключает возможность происхождения артефакта из иных кроме хапровской аллювиальной толщи геологических горизонтов.
В ходе анализа обнаружено два типа повреждений естественной формы metatarsus верблюда (рис. 4): сломы – отсутствуют проксимальный эпифиз, проксимальный конец диафиза, а также большая часть дистального эпифиза; следы рубки (рис. 4, а , справа) и резания (рис. 4, а , слева) на тыльной поверхности metatarsus, в дистальной части диафиза, по обеим его сторонам.
В ходе анализа ископаемой ко сти обнаружены два трасологически контрастных вида микрорельефа: общий рельеф фрагмента metatarsus и рельеф поверхностей сломов. Вся поверхно сть фрагмента metatarsus, кроме поверхностей сломов, покрыта
0 1 м 0 10 cм
а б
Рис. 3. Реконструкция древнего верблюда Paracamelus alutensis ( а ) и задняя правая метаподия животного ( б ). Художник Маурицио Антон.

Рис. 4. Дистальный фрагмент задней правой метаподии древнего верблюда Paracamelus alutensis (ЗИН № 35676) со следами резания-пиления ( а , с левой стороны) и рубки ( а , с правой).
пятнами коричнево-желтой и ржаво-железистой патины. Общий рельеф на микроуровне характеризуется незначительной окатанностью, сглаженностью и сплошной, мягко покрывающей все элементы микрорельефа заполировкой, которая напоминает покрытие лаком. При отно сительно небольших увеличениях (×10–20) нетрудно заметить, что заполировка блестящая (рис. 5). Характер заполировки свидетельствует о ее химическом происхождении (покрывает мельчайшие депрессии и каверны, во множестве присутствующие на микрорельефе) и появлении на уже видоизмененной минерализованной кости. Это обычный для костей из Ливенцовки вид химического выветривания, характерный не для костного вещества, а, скорее, для минералов. Поверх химической заполировки про слеживаются слабые следы более поздней заполировки механического происхождения, появившейся, видимо, в ходе анализа и камеральной обработки находки.
Микрорельеф поверхностей сломов иной. Он не имеет яркой заполировки и следов химического выветривания, более свежий на вид и не покрыт пятнами «железистой» патины.
Следы рубки и резания на ко сти образуют две группы. Левая группа (см. рис. 4, а ) состоит как минимум из семи следов пиления-резания. Каналы надрезов имеют V-образный профиль. Кинематика появления следов – скошенные по отношению к продольной оси ко сти разовые однонаправленные достаточно мощные надрезы (рис. 6). Правая группа представляет собой ряд как минимум из девяти следов рубки. Каналы зарубок в целом асимметричны, V-образные в сечении. Кинематика появления следов – скошенные по отношению к продольной оси кости, не сильные, практически прямые по отношению к поверхности рубящие удары (рис. 7). Надрубы широкие, в профиле U-образные только на самых выположенных участках

Рис. 5. Следы люстража на поверхности минерализованной кости, перекрывающие следы резания.
в центральной части канавок и V-образные – на концевых участках. Не имеет V-образного профиля только одна бороздка в центре группы.
Все бороздки несут признаки легкой окатанности (выступающие части микрорельефа оглажены), характерные для всей площади уцелевшей поверхности диафиза и метафиза данного обломка metatarsus. Самые тонкие и неглубокие канавки V-образного сечения, расположенные преимущественно на концах отдельных надрубов и/или надрезов, видимо, уничтожены окатанностью.
Погрызам хищников обычно соответствуют канавки U-образного профиля. Морфология этих повреждений зависит от размера пасти животного, характера захвата кости, формы и размеров последней. Такие погрызы сопровождаются отдельными точечными вдавлениями: зуб вдавливался в кость, не проскальзывая по ее поверхности и не оставляя U-образной канавки. Кроме того, следы зубов хищников имеют симметричный уплощенный профиль и располагаются на обеих сторонах кости (от верхней и нижней челюсти соответственно). В нашем случае оборотная сторона ко сти чистая, на ней нет никаких погрызов (см. рис. 4, б ). На боковых сторонах обломка metatarsus погрызы также отсутствуют (см. рис. 4, в ).
Описанные выше бороздки определяются нами как следы искусственного, не животного происхождения, т.к. они V-образные в сечении, не однообразные, являются результатом разных типов воздействия – пиления-резания и рубки. Иными словами, по локализации на кости, характеру распространения и нанесения, профилю ни одна из двух групп следов не может быть отнесена к погрызам хищников или грызунов.
Результаты экспериментов полностью подтверждают и уточняют эти выводы. Чоппером из окремнённого известняка (рис. 8) производились надрезы на свежей и сухой кости и ее рубка. Экспериментально

Рис. 7. Следы рубки на metatarsus из Ливенцовки.
Рис. 6. Следы пиления-резания на metatarsus из Ливенцовки.

Рис. 8. Чоппер из гальки окремненного известняка, использовавшийся в экспериментах по рубке и резанию кости.

Рис. 10. Экспериментальная зарубка, произведенная чоппером на сухой кости.

Рис. 9. Экспериментальные надрезы, произведенные по свежей кости чоппером из окремненного известняка.
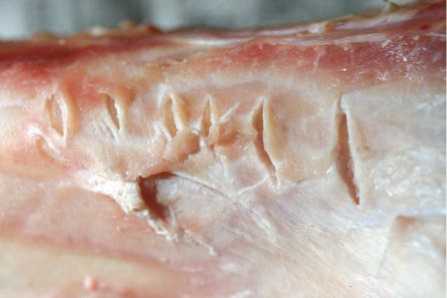
Рис. 11. Экспериментальные зарубки на поверхности неочищенной свежей кости.
полученные следы от пиления-резания (рис. 9) показали, что при резании свежей кости раздвоение канавок надрезов на их окончании (в начале надрезов гораздо реже) происходит не при повторном проходе лезвия по тому же месту, а при однократном однонаправленном движении из-за нерегулярности движения и изогнутости рабочего края орудия. Следы рубки сухой кости отличаются крупной выкрошенностью бортов канавок (рис. 10). Борта канавок надрубов, сделанных на свежей кости, имеют мелкую выкрошенность (рис. 11, 12).
Мелкая выкрошенность бортов канавок надрубов на описываемом нами образце metatarsus уничтожена окатанностью, однако общая морфология следов, отсутствие следов крупных фасеток выкрошенности свидетельствуют о том, что зарубки были сделаны на достаточно свежей кости. В ходе экспериментов с костями старых животных или просто сухими костями избежать появления такой выкрошенности практически не удается. Поэтому можно сделать вывод, что животное, которому принадлежала данная кость, вероятно, было относительно молодое, а кость относи-

Рис. 12. Следы рубки на поверхности свежей кости после чистки.
тельно свежая; обе группы следов появились во время разделки свежей туши животного.
Следы обеих групп, как следует из анализа общей морфологии, оставлены одним орудием с достаточно массивным и острым лезвием. Скорее всего, это был либо чоппер, либо массивный отщеп.
Плюсна верблюда практически целиком состоит из кожи, сухожилий и кости. Разделка в данном случае производилась не с целью отделения мясных частей от задней конечности. Изучаемые следы могли появиться при перерубании сухожилий для отделения мозолистого копыта верблюда. Характер следов и их расположение в нижней части тыльной поверхности metatarsus, в местах нахождения мощных сухожилий мышц – сгибателей пальцев стопы (m. flexor digitorum; рис. 13), соответствуют реконструируемой нами причине их появления. Для каких целей проводилось отделение копыт – не совсем ясно, возможно, для облегчения переноски более мясистых верхних частей конечности либо для отделения от кости самих сухожилий или даже снятия шкуры.
Артефакт из Ливенцовки является свидетельством того, что древний человек на территории нижнего Дона в борьбе за биоресурсы успешно конкурировал с обитавшими по соседству крупными хищниками: этрусским медведем Ursus cf. etruscus , гиенами Pliocrocuta perrieri и Pachicrocuta brevirostris , каракалом Lynx issidorensis , гепардом Acinonyx cf. pardinensis и крупной саблезубой кошкой Homotherium crenatidens .
Важно отметить, что результаты анализа данной находки представляют собой успешный, хотя и редкий пример использования трасологических данных для доказательства неприродного происхождения древнейших артефактов. Ранее трасологами были обнаружены следы разделки на костях вымерших африканских копытных возрастом 2,5 млн лет [Heinzelin et al., 1999]. И форма изделий, и следы рубки и резания являются равнозначными и равноправными источниками информации; они в одинаковой мере могут быть использованы для решения основных задач археоло-
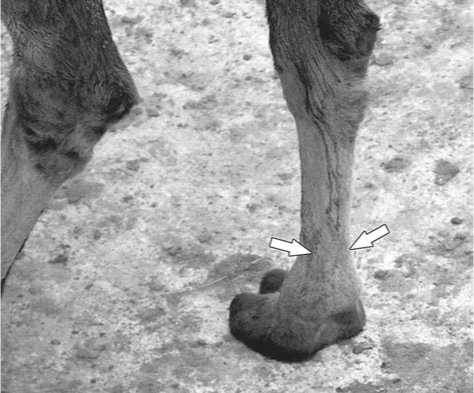
Рис. 13. Место расположения сухожилий мышц – сгибателей пальцев стопы (m. flexor digitorum) на плюсне современного верблюда.
гии как единственной научной дисциплины, способной по материальным остаткам реконструировать деятельность человека в далеком прошлом.
Контекст обнаружения кости древнего верблюда Paracamelus alutensis , степень ее сохранности и выразительность следов работы каменным орудием на поверхности находки, с нашей точки зрения, могут указывать на достаточно раннее появление человека в Евразии.
