Оценка динамики генетических процессов в молочном скотоводстве

Автор: Шендаков А.И.
Журнал: Биология в сельском хозяйстве @biology-in-agriculture
Рубрика: Актуальные вопросы отраслей животноводства
Статья в выпуске: 1 т.6, 2015 года.
Бесплатный доступ
Для решения проблемы были проведены исследования динамики генетических процессов в хозяйствах Орловской области. Все исследования проводились на чёрно-пёстрых коровах. Были изучены коэффициенты наследуемости и коэффициенты генетической корреляции между селекционными признаками, а также динамика этих параметров, коррелятивные сдвиги, эффекты селекции, паратипические корреляции, был предложен и апробирован метод вычисления множественной плейотропии признаков молочной продуктивности и пр. Выявлено существенное влияние на генотипическую изменчивость и генетические корреляции способов и интенсивности отбора, а также влияние инбридинга на наследуемость признаков молочной продуктивности. Проведённые исследования позволили сделать вывод, что в стадах чёрно-пёстрого скота при усилении интенсивности отбора (i=0,77-0,84σ) аддитивная изменчивость признаков молочной продуктивности может не только уменьшаться, но и возрастать. При этом до 50% поголовья имело положительную генетическую корреляцию между удоем и жирностью молока (r G=0,375) благодаря высокому аддитивному наследованию данных селекционных признаков, а при оптимальных вариантах выбраковки их генетическая корреляция равнялась 0,106-0,290 (р
Елекция, чёрно-пёстрый скот, отбор, генотипическая изменчивость, генотипическая и паратипическая корреляция, плейотропия
Короткий адрес: https://sciup.org/14770291
IDR: 14770291 | УДК: 636.082.23(241-242):
Текст научной статьи Оценка динамики генетических процессов в молочном скотоводстве
Введение. В современной селекции существуют разные методы увеличения молочной продуктивности: скрещивание, гибридизация, разведение по линиям и пр., но, так или иначе, успех применения данных методов зависит от множества факторов, одними из которых являются правильно организованный отбор [19] и подбор [5, 12, 21]. При этом предлагаются разные подходы при составлении родительских пар, в том числе подбор родителей по ЕАВ -локусу групп крови [8]. Как правило, аутбридинг и инбридинг дают разные результаты в селекции [16], а при разных степенях инбридинга результаты могут быть не только отрицательны, но и противоречивы [1, 3, 8, 13], инбридинг может влиять на соотношение полов в потомстве [17, 20, 27, 30] и, возможно, способствовать генетическим явлениям, которые при скрещивании и гибридизации принято называть эффектом гетерозиса. В связи с активным использованием искусственного осеменения в современном скотоводстве большинства развитых стран изучению проблем инбридинга и способов минимизации инбред-ной депрессии посвящено много научных работ [24, 27, 31, 32, 34, 35].
Возникает необходимость более подробного исследования генетических корреляций между селекционными признаками [25, 28, 30, 33], в том числе изучена генетическая корреляция между признаками бурого швицкого [28], голштинского [33], симментальского и чёрно-пёстрого скота [20]. В настоящее время ведётся модернизация и оптимизация селекционных программ [9, 18], а также программ разведения [6, 7], в том числе изучено иммуногенетическое сходствo молочных пород, наиболее распространенных в РФ. Совершенствуются методы разведения пород скота [15].
Однако следует признать, что в молочном скотоводстве постсоветского пространства оценке динамики генетических процессов при разных способах и формах отбора, а также при разных видах подбора родительских пар уделяется мало внимания, особенно мало внимания уделяется изучению генетикостатистических параметров при разных вариантах подбора, структуры генетической изменчивости и величины генетических корреляций не только удоев и жирности молока, но и других селекционных признаков. Моделированию селекционных процессов вообще не уделяется внимания, за исключением научных работ В. М. Кузнецова [8, 9]. Многие учёные, между тем, по-разному видят решения проблем отбора. В частности, И. М. Стародумов с соавт. (2007) предлагает ведение модального отбора и утверждает, что при разведении чёрно-пёстрого скота следует отдавать предпочтение коровам, у которых сумма нормированных отклонений по основным промерам тела соответствует показателям, средним в популяции, или несколько уступает им [14]. По данным М. С. Габаева с соавт. (2013), в стадах может проявляться скрытый стабилизирующий отбор, а его действие в определённой степени может нести характер дизруптивного отбора [4]. В племенных стадах отбор всегда сочетается с подбором, который, в зависимости от гетерогенности, может по-разному влиять на проявление аддитивной и неаддитивной изменчивости [21]. В научной статье В. Мымрин (2006) отмечает, что при увеличении продуктивности матерей коров по наивысшей лактации возрастает продуктивность дочерей по первой и второй лактациям [12].
При этом в каждом стаде существуют свои особенности взаимосвязи между признаками. Согласно исследованиям многих учёных, на первых этапах селекционной работы для быстрой оценки предрасположенности стада к одновременному увеличению признаков молочной продуктивности целесообразно использовать генетическую корреляцию (rG) и аддитивную изменчивость [11, 30]. Согласно современным представлениям о генетической корреляции между селекционными признаками, следует отметить два предположения: во-первых, взаимосвязь признаков обеспечивается аддитивным (т. е. суммирующим) действием генов; во-вторых, явление плейотропии способствует положительной корреляции между двумя признаками [2, 11].
Однако следует признать, что далеко не все признаки коррелируют положительно, а следовательно, не исключено, что в случае проявления отрицательной генетической корреляции в популяциях проявляются неаддитивные генетические эффекты, в том числе эпистаз, регрессия, доминирование или сверхдоминирование.
Так, согласно новым данным (Schrooten C., 2004), процентное содержание жира и белка в молоке кодируют гены, расположенные на 6 хромосоме, однако есть плейотропные гены, отвечающие за эти признаки (на 6 и 14 хромосомах), одновременное увеличение % белка и молочного белка в кг (на 6 хромосоме), одновременное увеличение % жира в молоке и количества молочного белка (на 23 хромосоме) и пр. [29]. В литературе также встречается информация о том, что плейотропное действие генов определяет живую массу телят во все периоды выращивания, однако не исключено, что помимо данного явления существенное влияние на рост и развитие оказывают аддитивные гены, и лишь часть из них проявляет плейотропию относительно к другим селекционным признакам. Схожая ситуация прослеживается по многим селекционным признакам (промерам вымени, признакам молочной продуктивности, скорости молокоотдачи, интервалам между отёлами и пр.) [29].
Говоря об этом, особое внимание следует обратить на работы зарубежных авторов, которые, в частности, выделяют не только аддитивное действие генов, но и аддитивно-доминантное, доминантное положительное и доминантное отрицательное действие аллелей и локусов. Аддитивную и неаддитивную генетическую изменчивость, по мнению A. M. Kelly, etc. (2009), в связи с возможностью их вычисления необходимо применять в селекционных программах, а в исследованиях A. Finley, S. Danerjee, P. Waldmann, T. Ericsson (2009) аддитивная изменчивость (h2) составляла 0,55, неаддитивная (d2) – 0,22, хотя при прогнозе составляла 0,60 и 0,13 соответственно [26]. В журнале «BMC Evolutionary Biology» приводится статья T. Bilde, etc. (2008), где авторы выделяют ядерную аддитивную изменчивость (σ2n), ядерную взаимодействующую изменчивость, включающую совместное дей- ствие аддитивных эффектов, доминирования и эпистаза (σ2t), материнский эффект (σ2m), отцовский эффект (σ2p), генетический эффект взаимодействия матерей и отцов (σ2k), изменчивость, обеспеченную взаимодействием внутрилинейных комбинаций (σ2rep), изменчивость между самками внутри кроссов линий (σ2w) [22].
Не менее интересны наработки зарубежных учёных в области растениеводства. Для примера можно отметить, что, по данным J.K. Conner, etc. (2002), у растений при выращивании в полевых условиях аддитивная генетическая изменчивость составляла 20-60% от аддитивной генетической изменчивости растений, выращенных в теплицах, а следовательно, согласно этим данным, улучшение условий возделывания сельскохозяйственных культур способствовало повышению эффективности селекции [23]. Известно также, что улучшение кормления и содержания может способствовать повышению уровня реализации генетического потенциала крупного рогатого скота, а в лучших условиях возрастает коэффициент наследуемости признаков молочной продуктивности [30].
Известно также, что в структуре генотипической изменчивости селекционных признаков доминантная и эпистатическая изменчивость составляют около 5-10%
[20, 30], а сам прогноз неаддитивного действия генов затруднителен. По нашему мнению, вопрос структуры генотипической изменчивости при дополнении анализом структуры фенотипических корреляций селекционных признаков приобретает более актуальное значение, поскольку в этом случае затрагивается ещё и вопрос паратипических корреляций. Множественные корреляции селекционных признаков (т. е., по Г. Ф. Лакину [10], корреляции трёх признаков) позволяют, на наш взгляд, говорить о сложных генетических процессах, обретающих фенотипическое выражение, а также о множественной плейотропии, т. е. о тех явлениях, когда один ген прямо или косвенно кодирует три и более признака, поскольку известно, что все живые организмы являются сложнейшими многофункциональными системами и все физиологические, биохимические и прочие процессы в организме взаимосвязаны (это предположение также делает А. В. Бакай с соавт, 2007 [2]). Из этого следует, что современная генетика, в связи недостатком научных данных, пока не может абсолютно точно ответить на вопрос о том, сколько генов отвечает за тот или иной количественный признак и какая их доля проявляет плейотропное действие, в том числе множественное плейотропное действие.
Данные факты объясняют актуальность наших исследований, целью которых был анализ динамики генетических процессов при отборе и составлении родительских пар в селекции молочного скота. В соответствии с целью исследований были поставлены основные задачи изучения следующих величин и процессов: 1) динамики аддитивной генотипической изменчивости удоев, генетических корреляций между удоем и жирностью молока в стаде коров при одностороннем и искусственно стабилизируемом отборе, а также отборе по независимым уровням выбраковки; 2) генетикостатистических параметров при одностороннем отборе по удою, в том числе селекционных эффектов, генотипических и паратипических корреляций между признаками молочной продуктивности, множественного плейотропного действия генов, кодирующих удой, на жирность молока и количество молочного жира; 3) распределения превосходства дочерей над матерями по жирности молока; структуры генетической изменчивости жирности молока и результатов увеличения молочной продуктивности чёрно-пёстрых коров, полученных при разной степени гетерогенности подбора и пр.
Материалы и методы исследований
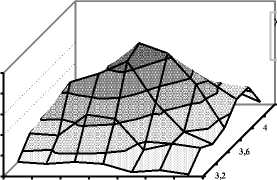
Моделирование отбора было проведено в ОПХ «Стрелецкое» при ВНИИ зернобобовых культур и ЗАО «Куракинское» Орловской области. Дойные стада хозяйств были представлены коровами с кровностью от 12,5 до 87,5% по голштинской породе. В стаде ОПХ «Стрелецкое» по всему поголовью прослеживалось нормальное распределение не только по удоям, но и по жирности молока: в целом, удой варьировал в пределах от 1500 до 7500 кг молока от одной коровы, однако 90% коров показывали удои от 3000 до 6000 кг молока (см. рис. 1). При удоях 4500 кг молока 47 коров имели жирность молока 3,80%. Нормальное распределение по удоям проявлялось и в ЗАО «Куракинское».
Голов, n
4,4
ирность молока,
%
3000 3500 4000 4500 5000 5500 6000
Удой за 305 дней, кг
40-50
30-40
20-30
10-20
0-10
Рис. 1. Распределение чёрно-пёстрых коров по удою и жирности молока в ОПХ «Стрелецкое» Орловской области
По методике Г. Ф. Лакина (1991) были изучены множественные корреляции между селекционными признаками rx (У, z)
' r 2xy + r 2xz - 2 r xyrzzryz
1 - r 2 yz где x, y и z
– взаимосвязанные признаки [10]. Генетическая корреляция между промерами и удоем, удоем и содержанием жира в молоке определялась по фор- мулам Хейзеля:
rG
I r x g У м " r y g x M (1)
V rxgxM " ГУ»Ум или _ ( rxgУ м rG = I—
+ ry g xм ):2 (2) [11], где фенотипические
■r x д x м y д y м
коэффициенты корреляций (r) можно выразить графически (см. рис. 2).
Дочери (удой) х y (живая масса)

Матери (удой) x y (живая масса)
Рис. 2. Фенотипические корреляции при вычислении генетической корреляции между удоем и живой массой (rG)
При этом учитывалось, что наличие нереального коэффициента наследуемости даже по одному из признаков, учитываемых в формуле, не даёт возможности вычисления генетической корреляции, а при наличии одного отрицательного коэффициента в числителе использовалась вторая формула Хейзеля. Селекционные дифференциалы удоев ( Sd ) рассчитывались как разница средних величин между группой отобранных коров и общего поголовья стада. Эффекты селекции были определены по классической формуле SE=h2·Sd , где h2 – коэффициент наследуемости, или доля генетической изменчивости в общей фенотипической вариации признака. Интенсивность отбора по удою (i) выражалась в долях σ, вычисленной для каждого стада в отдельности. Коррелятивные сдвиги селекционных признаков были определены по методике академика Л. С. Жебровского:
R _ SEx ■ hy ■ ay , где SEX - эффект селекции по призна- y = hx • ax rG ку х, hx и hy – детерминация фенотипа генотипом по признакам x и y (-Jh^), a - стандартное отклонение признаков x и y, rG – генетическая корреляция. Дисперсионный анализ проводили с определением критерия Краскелла-Уоллиса H=(Σ(Σxi))2:N, общей девиаты Dy=Σ(Σxi2)-H, факториальной девиаты Dx=Σ(Σxi)2:n-H, силы влияния h2x=Dx/Dy (с использованием F-критерия Фишера). Для определения динамики аддитивной изменчивости (h2) в стаде ОПХ «Стрелецкое» применялись уравнения линейной и нелинейной регрессии, в том числе уравнения, имеющие математическое выражение гипербол первого порядка: b .
y x a +
x
При отборе по молочной продуктивности в качест- ве одного из вариантов применялся индекс: , ПУДуд - Пс . h П% Ж - Пс Пж - Пс , где Тот. - индекс
I om. = b l + b 2 + b 3
a a 2 a 3
отбора; П – продуктивность коровы по удою, процентному содержанию и количеству молочного жира, ПС – средние значения признаков в стаде; b – весовые коэффициенты, определяемые по соответствующим признакам отбора на основе общей теории построения селекционных индексов; σ – стандартные отклонения по учтённым признакам. Также были определены селекционные эффекты по удою при разных вариантах отбора, аддитивная генетическая изменчивость удоев при разных способах и вариантах отбора, экономическая эффективность, прибыль от разных моделей отбора и пр.
Для вычисления коэффициентов паратипической корреляции применялась формула:
r f ; p 2 - h 1 • h 2 • rA, A 2 , где r - паратипическая кор-
EE
2
=
Ti
r
h
^;
•
реляция; r – фенотипическая корреляция; r – генетическая корреляция ( r G ), вычисляется по формулам Хейзеля; h2 – коэффициенты наследуемости соответствующих признаков; h – детерминация фенотипа генотипом по соответствующему признаку, т. е. корень квадратный из коэффициента наследуемости: T h 2 (при этом коэффициент наследуемости не возводился в степень, h2 являлся обозначением, следующим из традиции, заложенной Райтом и другими учёными, так же, как в формуле Л. С. Жебровского).
В стаде ОПХ «Стрелецкое» было вычислено влияние интенсивности одностороннего отбора по удою (от 10 до 90% выбраковки) на генетикостатистические параметры, в том числе на динамику коэффициентов вариации (C v ), селекционных дифференциалов (Sd), селекционных эффектов (SE), коэффициентов наследуемости (h2), детерминации фенотипа генотипом (h) и пр., а также на структуру фенотипических корреляций и множественный плейотропный эффект (удой – % жира в молоке – количество молочного жира). Множественный плейотропный эффект вычислялся на основе преобразованной множественной корреляции Г. Ф. Лакина (1991). При этом в качестве частных коэффициентов корреляций в формуле использовались частные коэффициенты генетической корреляции Хейзеля, при построении формул для вычисления и анализе с целью выявления генов, кодирующих соответствующие признаки, использовались три комбинации: , r и r .
G 1 ( G 2 G 3) G 2 ( G 1 G 3 ) G 3( G 1 G 2 )
При сравнении степени детерминации селекционными признаками матерей матерей аналогичных признаков у матерей и дочерей (пробандов) в аутбредной и инбредной группах стада чёрнопёстрых голштинизированных коров СПК им. Мичурина Верховского района Орловской области применялась множественная корреляция r . Аутбредные и инбредные коровы-первотёлки являлись аналогами по возрасту. Статистическая обработка данных и моделирование отбора проходили в компьютерной программе «Microsoft Excel».
Результаты и их обсуждение.
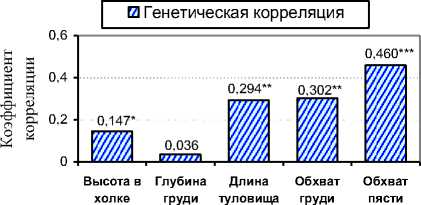
Ряд исследований, проведенных нами, позволяет подчеркнуть важные закономерности, выявленные в популяции молочного скота Орловской области. Так, между всеми изученными промерами у чёрно-пёстрых высокопродуктивных коров племенных организаций прослеживалась положительная генетическая связь – от 0,062 до 0,534. Удой генетически был связан с основными промерами: r G =0,036-0,460 (см. рис. 3). Наибольшая множественная корреляция между основными промерами была обнаружена у быков-производителей отечественного и европейского происхождения: rx ( y,z ) =0,609-0,872 и 0,647-0,941 соответственно.
Промеры
Рис. 3. Генетическая корреляция между удоем и основными промерами у чёрно-пёстрых коров, r G
При отборе по живой массе возрастание интенсивности выбраковки полновозрастных коров до 90% сопровождалось увеличением селекционного дифференциала и эффекта селекции по высоте в холке, глубине груди и обхвату груди (см. таблицу 1). При этом селекционный эффект (SE) по длине туловища не имел чёткой тенденции к увеличению – от ≈0,23 до 3,43 см, вы- сота в холке увеличивалась на ≈6,07 см, обхват груди – на ≈10,63 см.
При возрастании интенсивности отбора по высоте в холке от 10 до 90% коррелятивный сдвиг по глубине груди увеличивался от 0,33 до 2,70 см. Аналогично при положительной генетической корреляции (rG) отбор по одному из промеров вел к увеличению других промеров.
Таблица 1 – Селекционный эффект по промерам при отборе чёрно-пёстрых коров по живой массе (n=150)
|
% выбраковки |
Промеры, см |
|||||||
|
Высота в холке |
Глубина груди |
Длина туловища |
Обхват груди |
|||||
|
Sd |
SE |
Sd |
SE |
Sd |
SE |
Sd |
SE |
|
|
10 |
0,710 |
0,534 |
0,486 |
0,338 |
0,690 |
0,229 |
0,500 |
0,443 |
|
20 |
1,820 |
1,369 |
0,617 |
0,429 |
1,250 |
0,415 |
1,000 |
0,886 |
|
30 |
2,567 |
1,930 |
2,200 |
1,531 |
3,433 |
1,140 |
2,300 |
2,038 |
|
40 |
2,956 |
2,223 |
2,200 |
1,531 |
3,441 |
2,395 |
2,215 |
0,735 |
|
50 |
2,959 |
2,225 |
2,200 |
1,531 |
3,444 |
1,143 |
2,222 |
1,969 |
|
60 |
4,398 |
3,308 |
2,700 |
1,879 |
4,666 |
1,549 |
5,830 |
5,165 |
|
70 |
4,870 |
3,662 |
2,600 |
1,810 |
5,133 |
1,704 |
7,400 |
6,556 |
|
80 |
6,067 |
4,562 |
3,867 |
2,691 |
5,000 |
1,660 |
8,333 |
7,383 |
|
90 |
8,070 |
6,069 |
5,200 |
3,619 |
10,33 |
3,430 |
12,00 |
10,630 |
Из анализа таблицы 2 следует, что с возрастанием интенсивности одностороннего отбора по удою до 20%, или 0,39σ, в стаде ОПХ «Стрелецкое» наблюдалось уменьшение аддитивной генетической изменчивости удоя за 305 дней лактации и количества молочного жира – с 36,2 и 31,0% до 20,4 и 17,0% соответственно. При возрастании интенсивности до 60%, или 0,96σ, аддитивная генетическая изменчивость по удою стабильно понижалась до 3,4%, по количеству молочного жира – до 6,3%, в то время как при выбраковке с интенсивностью 0,84σ генетическая изменчивость жирности молока возросла до 22,8%, количества молочного жира – до 15,8%. Это подтверждало возможности отбора, однако прогноз удоев дочерей через Sd·h2 дал результаты, превышающие их фактические удои на 33-124 кг молока и 1,4-2,8 кг молочного жира при интенсивности 0,23-0,69σ. Данное обстоятельство даёт возможность сделать вывод о существенном влиянии паратипических условий на эффект отбора по селекционным признакам, имеющим высокие коэффициенты фенотипической изменчивости. Также можно сделать предположение о весьма удачных сочетаниях у 20-30% поголовья и проявлении неаддитивных генетических эффектов, полученных от отдельных быков-производителей, таких, как Риголетто 9862, Лель 23682 и Хезелден 474459. Не исключено, что в связи с высоким коэффициентом наследуемости в группах коров с кровностью 12,5, 25, 62,5 и 75% по голштинам проявлялась взаимное усиле- ние аддитивной и неаддитивной изменчивости. В то же время, коровы поколения F1 отличались низкой аддитивной изменчивостью и невысокими удоями, что, вероятно, связано с проявлением процесса адаптации помесей и неблагоприятным проявлением взаимосвязи «генотип-среда».
При отборе по удою за 305 дней при уровнях 2000 и 2500 кг молока интенсивность выбраковки составила всего 0,02σ и 0,10σ (SE = 6 и 25 кг молока соответственно). При ужесточении отбора и установлении порога в 3750 кг молока аддитивная генетическая изменчивость удоя и количества молочного жира понизилась до 6,0 и 5,0% соответственно, а при пороге 4000 кг молока (или 1,15σ) наблюдалось увеличение h2А жирности молока – до 30,2%. Тем не менее, данный порог был недопустим для практического применения, как и порог в 3500 кг молока, поскольку даже при нём интенсивность отбора составляла 0,72σ, или 45,6% выбраковки. Наиболее рациональным был порог 30003250 кг молока по первой лактации, что соответствовало 20-30% выбраковки из стада (0,35-0,49σ) и привело к селекционному эффекту (SE) в 89-125 кг молока при стабильных условиях кормления.
Таблица 3 – Влияние % генов голштинской породы на аддитивную изменчивость удоев у коров при разных уровнях выбраковки из стада ОПХ «Стрелецкое» Орловской области
|
Генотип, % HF |
h2 А удоев при разных уровнях выбраковки из стада, % |
||
|
Без выбраковки |
3500 кг |
4000 кг |
|
|
0 (контроль) |
1,0 |
55,8 |
9,4 |
|
25,0% |
30,2 |
27,4 |
– |
|
50,0% |
7,4 |
– |
– |
|
62,5% |
44,4 |
50,0 |
39,8 |
|
75,0% |
55,0 |
48,0 |
33,4 |
Таблица 2 – Влияние вариантов, способов и интенсивности отбора на аддитивную изменчивость и генетическую корреляцию селекционных признаков чёрно-пёстрого скота в ОПХ «Стрелецкое»
|
Вариант, способ отбора, % выбраковки, количество выбывших коров |
н О ••* § о я ® Н О к К |
Аддитивная изменчивость селекционных признаков, % |
Фенотипические корреляции между селекционными признаками молочной продуктивности, r±m r |
u И |
|||||
|
1^ |
Удой матерей -удой дочерей |
Жир матерей -жир дочерей |
Удой матерей -жир дочерей |
Жир матерей -удой дочерей |
|||||
|
Односторонний отбор по удою (n=542, σ=704 кг) |
|||||||||
|
Без выбраковки |
– |
36,2 |
16,0 |
31,0 |
0,181±0,029*** |
0,080±0,036* |
0,003±0,043 |
-0,010±0,044 |
-0,029 |
|
10% (n=54) |
0,23σ |
26,6 |
16,6 |
23,8 |
0,133±0,033*** |
0,083±0,037* |
0,011±0,044 |
-0,042±0,048 |
-0,148* |
|
20% (n=108) |
0,39σ |
20,4 |
13,2 |
17,0 |
0,102±0,039** |
0,066±0,042 |
-0,020±0,050 |
-0,057±0,054 |
0,411*** |
|
30% (n=163) |
0,54σ |
16,6 |
14,4 |
13,0 |
0,083±0,043 |
0,072±0,044 |
-0,007±0,052 |
-0,072±0,059 |
0,290** |
|
40% (n=217) |
0,69σ |
15,6 |
16,6 |
14,0 |
0,078±0,047 |
0,083±0,047 |
-0,024±0,058 |
-0,056±0,062 |
0,456*** |
|
50% (n=271) |
0,84σ |
7,8 |
22,8 |
15,8 |
0,039±0,056 |
0,114±0,048* |
-0,064±0,069 |
-0,049±0,067 |
0,840*** |
|
60% (n=325) |
0,96σ |
3,4 |
17,4 |
6,3 |
0,017±0,066 |
0,087±0,057 |
-0,005±0,069 |
-0,051±0,075 |
0,172* |
|
Отбор по удою с независимыми уровнями выбраковки, (n=542, σ=704 кг) |
|||||||||
|
2000 кг (n=4) |
0,02σ |
33,4 |
14,8 |
29,2 |
0,167±0,030*** |
0,074±0,037* |
0,011±0,042 |
-0,017±0,045 |
-0,027 |
|
2500 кг (n=29) |
0,10σ |
32,2 |
14,2 |
26,6 |
0,161±0,031*** |
0,071±0,038 |
-0,010±0,045 |
-0,036±0,047 |
0,177* |
|
3000 кг (n=110) |
0,35σ |
19,8 |
6,7 |
16,6 |
0,099±0,039* |
0,067±0,042 |
-0,020±0,050 |
-0,052±0,053 |
0,396*** |
|
3250 кг (n=162) |
0,49σ |
17,4 |
13,4 |
13,8 |
0,087±0,043 |
0,073±0,044 |
-0,001±0,051 |
-0,071±0,059 |
0,106** |
|
3500 кг (n=246) |
0,72σ |
11,6 |
19,2 |
12,4 |
0,058±0,052 |
0,096±0,048* |
-0,052±0,064 |
-0,042±0,063 |
0,627*** |
|
3750 кг (n=306) |
0,93σ |
6,0 |
17,0 |
5,0 |
0,030±0,061 |
0,085±0,055 |
-0,007±0,066 |
-0,070±0,075 |
0,192* |
|
4000 кг (n=368) |
1,15σ |
0,6 |
30,2 |
0,5 |
0,003±0,076 |
0,151±0,055** |
-0,001±0,076 |
-0,005±0,077 |
0,011 |
|
Искусственный стабилизирующий отбор по удою (n=542, σ=704 кг) |
|||||||||
|
2000-5500 кг |
0,007σ |
36,4 |
15,0 |
32,0 |
0,182±0,029*** |
0,075±0,037* |
0,024±0,041 |
-0,019±0,045 |
0,021 |
|
2300-5200 кг |
0,010σ |
36,4 |
12,8 |
30,8 |
0,182±0,029*** |
0,064±0,038 |
0,006±0,043 |
-0,024±0,046 |
-0,108* |
|
2600-4900 кг |
0,049σ |
37,4 |
14,6 |
31,0 |
0,187±0,030*** |
0,073±0,039 |
-0,018±0,047 |
-0,038±0,049 |
0,234** |
|
2900-4600 кг |
0,099σ |
31,6 |
8,2 |
27,2 |
0,158±0,035*** |
0,041±0,045 |
0,009±0,048 |
-0,050±0,054 |
-0,256** |
|
3200-4300 кг |
0,139σ |
24,0 |
8,2 |
20,6 |
0,120±0,044** |
0,041±0,053 |
0,094±0,047* |
-0,068±0,065 |
0,186* |
|
3500-4000 кг |
0,166σ |
13,0 |
6,4 |
10,8 |
0,065±0,078 |
0,032±0,083 |
0,072±0,078 |
-0,113±0,110 |
-0,446*** |
Примечание: достоверность коэффициентов корреляции: *– p<0,05, **– p<0,01, ***– p<0,001 (для всех таблиц и рисунков).
При этом пороги в 3500 и 4000 кг молока (см. таблицу 3) дали неплохие результаты по аддитивной генетической изменчивости удоев у коров с 62,5 и 75% генов голштинской породы – от 50,0 до 39,8% и от 48,0 до 33,4% соответственно, а следовательно, для высококровного по голштинам скота целесообразны более высокие уровни выбраковки по первой лактации при жёстком отборе – 4000 кг молока и выше. Отбор полукровных коров был малоэффективным.
Как правило, стабилизирующая форма отбора прослеживается только в естественных условиях обитания, когда наиболее приспособленными к существованию являются особи со средними значениями признака в популяции. В молочном скотоводстве стабилизирующий отбор применяют при совершенствовании экстерьера, однако в стадах с интенсивным использованием дойных коров нередко возникают условия, подоб- ные тем, которые складываются в диких популяциях, т. е. возрастает значимость «выживания» животных, коровы с высокими удоями чаще заболевают маститами, туберкулёзом, лейкозом и пр. болезнями, в результате чего при несвоевременном ветеринарном вмешательстве могут выбывать из стада. Моделирование стабилизирующего отбора в ОПХ «Стрелецкое» показало, что при нём аддитивная генетическая изменчивость уменьшалась слабее и несущественно возрастала интенсивность отбора – от 0,007σ до 0,166σ. Так, даже при формировании для воспроизводства стада с удоями от 3500 до 4000 кг молока по первой лактации h2А удоя, жирности молока и количества молочного жира составил 13,0, 6,4 и 10,8% соответственно.
В стаде ОПХ «Стрелецкое» 542 коровы-первотёлки имели слабую отрицательную генетическую корреляцию между удоем и жирностью молока (rG=-0,029). При одностороннем отборе по удою только при выбраковке 10% (0,23σ) была получена отрицательная генетическая корреляция (rG=-0,148). При возрастании интенсивности отбора до 0,84σ наблюдался рост данного показателя до 0,840, что, очевидно, можно преимущественно объяснить возрастанием аддитивной изменчивости жирности молока. Некоторое уменьшение rG при 30%, или i=0,54σ, очевидно, можно объяснить тем, что в группу, оставленную для воспроизводства, входили дочери, полученные от быков с низким потенциалом жирности молока. Однако фенотипическая корреляция между удоем и жирностью мо- лока у коров-дочерей при всех изученных вариантах интенсивности выбраковки составила 0,020-0,085.
Аналогичные тенденции по величине генетической корреляции были получены при выбраковке по установленным уровням. Так, отрицательный коэффициент r G был получен только при выбраковке коров с продуктивностью ниже 2000 кг молока (-0,027). При интенсивности 0,72σ, или уровне 3500 кг, была получена максимальная генетическая корреляция при данном способе отбора (rG=0,627). В данном случае фенотипическая корреляция между удоем и жирностью молока варьировала от 0,027 до 0,117, т. е. была слабо положительной.
При моделировании стабилизирующего отбора было получено чередование положительных и отрицательных коэффициентов генетической корреляции: при
0,010σ, 0,099σ и 0,166σ rG составил -0,108, -0,256 и -0,446 соответственно, проявляя чёткую тенденцию к ослаблению связи. При остальных вариантах стабилизации признака генетическая корреляция составила от 0,021 до 0,234.
В целом, всё способы отбора подтвердили возможность получения более благоприятных генетических корреляций при его моделировании, однако в стаде аддитивная генетическая изменчивость удоев не детерминировала генетическую корреляцию между этим признаком и жирностью молока, в то время как аддитивная генетическая изменчивость жирности молока определяла r G на r2=21,1%. Это позволяет сделать вывод о большем значении величины жирности молока для формирования корреляционной связи.
Таблица 4 – Влияние вариантов, способов и интенсивности отбора на аддитивную изменчивость и генетическую корреляцию селекционных признаков чёрно-пёстрого скота в ЗАО «Куракинское»
|
Вариант, способ отбора, % выбраковки, количество выбывших коров |
н о S S g ° к |
Аддитивная изменчивость селекционных признаков, % |
Фенотипические корреляции между селекционными признаками молочной продуктивности, r±m r |
5 ° £ S |
|||||
|
g> |
Удой матерей -удой дочерей |
Жир матерей -жир дочерей |
Удой матерей -жир дочерей |
Жир матерей -удой дочерей |
|||||
|
Односторонний отбор по удою (n=215, σ=808 кг) |
|||||||||
|
Без выбраковки |
– |
14,5 |
– |
10,2 |
0,072±0,068 |
-0,060±0,068 |
-0,011±0,068 |
-0,049±0,068 |
– |
|
10% (n=21) |
0,219σ |
27,4 |
– |
16,6 |
0,137±0,071 |
-0,054±0,072 |
-0,024±0,072 |
-0,117±0,071 |
– |
|
20% (n=42) |
0,370σ |
33,8 |
– |
20,2 |
0,169±0,074* |
-0,033±0,076 |
-0,025±0,076 |
-0,144±0,075 |
– |
|
30% (n=64) |
0,497σ |
30,8 |
– |
20,3 |
0,154±0,080 |
-0,041±0,081 |
0,006±0,082 |
-0,123±0,080 |
– |
|
40% (n=85) |
0,636σ |
35,8 |
– |
25,6 |
0,179±0,085* |
-0,019±0,088 |
0,016±0,088 |
-0,108±0,087 |
– |
|
50% (n=107) |
0,768σ |
31,2 |
1,4 |
25,2 |
0,158±0,094 |
0,007±0,097 |
-0,003±0,097 |
-0,047±0,096 |
0,357* |
|
60% (n=128) |
0,899σ |
25,3 |
2,2 |
24,1 |
0,126±0,108 |
0,011±0,108 |
-0,007±0,108 |
-0,002±0,108 |
0,010 |
|
Отбор по удою с независимыми уровнями выбраковки (n=215, σ=808 кг) |
|||||||||
|
2000 кг (n=2) |
0,032σ |
16,8 |
– |
11,6 |
0,084±0,068 |
-0,061±0,068 |
-0,061±0,068 |
-0,002±0,069 |
– |
|
2500 кг (n=5) |
0,069σ |
19,6 |
– |
13,0 |
0,098±0,068 |
-0,056±0,070 |
-0,074±0,069 |
-0,008±0,069 |
– |
|
3000 кг (n=8) |
0,169σ |
26,2 |
– |
16,8 |
0,131±0,068 |
-0,061±0,069 |
-0,101±0,069 |
-0,021±0,070 |
– |
|
3250 кг (n=10) |
0,188σ |
26,8 |
– |
17,2 |
0,134±0,069 |
-0,062±0,070 |
-0,104±0,069 |
-0,019±0,070 |
– |
|
3500 кг (n=16) |
0,247σ |
28,6 |
– |
16,8 |
0,143±0,070* |
-0,056±0,071 |
-0,134±0,070 |
-0,018±0,071 |
– |
|
3750 кг (n=26) |
0,313σ |
30,6 |
– |
17,4 |
0,153±0,071* |
-0,041±0,073 |
-0,154±0,071* |
-0,020±0,073 |
– |
|
4000 кг (n=42) |
0,438σ |
32,8 |
– |
23,0 |
0,164±0,074* |
-0,032±0,076 |
-0,103±0,075 |
-0,007±0,076 |
– |
|
Искусственный стабилизирующий отбор по удою (n=215, σ=808 кг) |
|||||||||
|
2000-5500 кг |
-0,130σ |
11,6 |
– |
9,7 |
0,058±0,071 |
-0,054±0,072 |
-0,009±0,072 |
-0,004±0,072 |
– |
|
2300-5200 кг |
-0,195σ |
– |
– |
– |
-0,010±0,074 |
-0,053±0,074 |
-0,027±0,074 |
-0,016±0,074 |
– |
|
2600-4900 кг |
-0,374σ |
– |
– |
– |
-0,064±0,083 |
-0,044±0,083 |
0,008±0,083 |
0,004±0,083 |
– |
|
2900-4600 кг |
-0,498σ |
– |
– |
– |
-0,043±0,094 |
-0,053±0,094 |
0,025±0,094 |
-0,044±0,094 |
– |
|
3200-4300 кг |
-0,631σ |
– |
– |
– |
-0,019±0,121 |
-0,110±0,120 |
0,038±0,121 |
-0,047±0,121 |
– |
|
3500-4000 кг |
-0,776σ |
– |
– |
– |
-0,041±0,189 |
-0,210±0,185 |
0,195±0,185 |
-0,134±0,187 |
– |
Из таблицы 4 следует, что в ЗАО «Куракинское» с усилением интенсивности выбраковки по удою до 4050%, или до 0,636-0,768σ, проявлялась тенденция к возрастанию аддитивной генетической изменчивости по удою и количеству молочного жира от 14,5 и 10,2 до 35,8-31,2 и 25,6-25,2% соответственно. Невысокие коэффициенты наследуемости жирности молока были получены только при выбраковке 50 и 60% (h2А =1,4 и 2,2% соответственно). Аналогично только при этих вариантах выбраковки была получена положительная генетическая корреляция между удоем и жирностью молока: rG=0,357 и 0,010, в то время как в других вариантах она не была рассчитана из-за отсутствия аддитивного наследования процентного содержания жира в молоке. При выбраковке по удою с независимыми уровнями прослеживались тенденции, схожие с односторонним отбором по удою, однако в этом случае даже при пороге в 4000 кг молока (i=0,438σ) не было получено аддитивной генетической изменчивости жирности молока и положительной генетической корреляции между удоем и жирностью молока у коров. Искусственный стабилизирующий отбор не дал существенных результатов и был нецелесообразен.
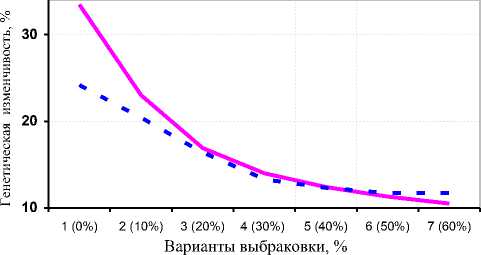
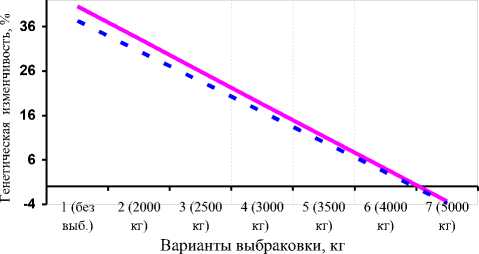
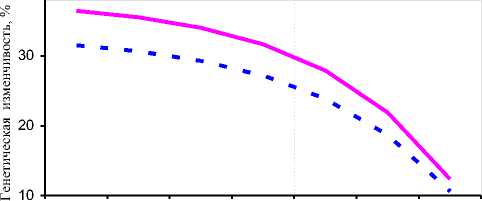
Полученные результаты показали, что при чётком нормальном распределении коров по удоям и жирности молока в ОПХ «Стрелецкое» спад аддитивной генетической изменчивости в связи с односторонним отбором по удою можно выразить урав- нениями гипербол 1-го порядка: у _ 5 683 + 33,483 - для xx удоев, где х – вариант выбраковки (1 – 0%, каждый следующий +10%); у = 8291 + 24,244 - для количест-xx ва молочного жира (см. рис. 4, А). При этом прогноз линий регрессии существенно коррелировал с фактическими значениями (r=0,942 и 0,943 соответственно). При отборе коров по удоям с независимыми уровнями выбраковки ослабление аддитивной генетической изменчивости удоев и количества молочного жира выражалось уравнениями линейной регрессии:
у, = 47,88 - 7,308 x и ух = 44,14 - 6,857 x соответственно (см. рис. 4, Б), где х – вариант выбраковки (1 – без выбраковки, каждый следующий +500 кг молока). Совпадение линейной регрессии с эмпирической кривой также подтвердилось высокими корреляциями (r=0,964-0,966). В отличие от одностороннего отбора по удою, при искусственном стабилизирующем отборе проявлялась тенденция первоначального сохранения и даже незначительного возрастания генетической изменчивости, а затем резкого спада. Такая закономерность была нами выражена формулами: у^ = 38,05 - 1,59 x — для удоев, ух = 33,05 - 1,56 x — для молочного жира (см. рис. 4, В), где х – вариант выбраковки (1 – без выбраковки, 2 – 3500 кг молока между крайними границами обора, каждый следующий – -600 кг молока). Корреляции между фактической кривой и графиком прогноза по уравнениям составили 0,975 и 0,980 соответственно.
Также следует заметить, что в ОПХ «Стрелецкое» при одностороннем отборе по удою не было обнаружено чётких тенденций в изменении аддитивной наследуемости процентного содержания жира в молоке. При отборе по удою с независимыми уровнями выбраковки проявлялась параболическая зависимость генетической изменчивости жирности молока, а при стабилизирующем отборе – линейный спад.
В отличие от ОПХ «Стрелецкое», в ЗАО «Куракин-ское» при одностороннем отборе по удою проявлялась параболическая зависимость его генетической изменчивости, которую, построив по схожему принципу, можно выразить уравнением параболы 2-го порядка: ух = 3,4 +14,167x -1,583x2, где х — вариант выбраковки (1 – 0%, каждый следующий +10%). Корреляция значений параболической кривой с эмпирическими значениями составила 0,934. Генетическая изменчивость количества молочного жира при этом возрастала по линейной регрессионной зависимости: ух = 18,167 + 2,586x (корреляция с фактическими показателями составила 0,956). При отборе по независимым уровням выбраковки также прослеживалось возрастание генетической изменчивости удоев и количества молочного жира, что выражалось линейными зависимостями. За исключением первого варианта выбраковки (размах границ, R=3500 кг молока), искусственно стабилизируемый отбор не дал аддитивной генетической изменчивости по признакам молочной продуктивности.
Удой, кг Молочный жир, кг
А) односторонний отбор по удою
Удой, кг Молочный жир, кг
Б) отбор по удою с независимыми уровнями выбраковки
Удой, кг Молочный жир, кг
1 (без выб.) 2 (R=3500 3 (R=2900 4 (R=2300 5 (R=1700 6 (R=1100 7 (R=500
Варианты выбраковки, кг
В) искусственный стабилизирующий отбор по удою Рис. 4. Динамика аддитивной изменчивости при разных вариантах отбора в ОПХ «Стрелецкое»
В ОПХ «Стрелецкое» разные варианты отбора с увеличением интенсивности выбраковки давали эффекты селекции от 48 до 245 кг, от 6 до 293 кг и от 2 до 45 кг молока. В ЗАО «Куракинское» положительные эффекты составляли от 26 до 149 кг и от 4 до 51 кг молока. Вместе с тем, в этом хозяйстве при моделировании стабилизирующего отбора в группе от 2900 до 4600 кг молока стало проявляться превосходство дочерей над матерями на 1% (см. таблицу 5), а дальнейшая стабилизация признака привела к улучшению на 8% при отрицательном эффекте селекции, согласно прогнозу (-91 кг молока). Это позволяет сделать вывод о том, что в среднестатистических и ухудшающихся условиях кормления были наиболее выносливы коровы с удоями от -500 до -700 кг от среднего значения в стаде. При этом были очевидны погрешности удоев, прогнозируемых через формулу SE=h2·Sd (1-7%), и существенные различия эффектов селекции в разных стадах при одинаковых вариантах и способах отбора. Дисперсионный анализ показал, что сила влияния фактора «стадо» на эффект селекции в данном случае составила 24,6±2,1% (α<1%), на отношение удоев дочерей к удоям матерей – 62,3±1,0% (α<1%), на достоверность про- гноза удоев по формуле SE=h2·Sd – 87,3±0,3% (α<1%). Следовательно, эффективность селекции в разных стадах может существенно различаться даже при одинаковых способах и формах отбора. В отдельных случаях стабилизирующий отбор может давать незначительный эффект селекции по удою (от 2 до 45 кг молока).
Таблица 5 – Эффективность селекции чёрно-пёстрого скота по удою и достоверность прогноза увеличения удоев при разных вариантах выбраковки в хозяйствах
|
Вариант, способ отбора, % выбраковки, количество выбывших коров |
ОПХ «Стрелецкое» |
ЗАО «Куракинское» |
||||
|
SE удоя |
Относительно к матерям, % |
Относительно к прогнозу, % |
SE удоя |
Относительно к матерям, % |
Относительно к прогнозу, % |
|
|
Односторонний отбор по удою |
||||||
|
10% (n=54 и 21) |
48 |
97 |
99 |
26 |
90 |
93 |
|
20% (n=108 и 42) |
88 |
95 |
99 |
43 |
88 |
93 |
|
30% (n=163 и 64) |
126 |
93 |
98 |
58 |
87 |
93 |
|
40% (n=217 и 85) |
163 |
91 |
97 |
74 |
85 |
93 |
|
50% (n=271 и 107) |
203 |
90 |
97 |
90 |
83 |
93 |
|
60% (n=325 и 128) |
245 |
87 |
97 |
149 |
95 |
94 |
|
Отбор по удою с независимыми уровнями выбраковки |
||||||
|
2000 кг (n=4 и 2) |
6 |
99 |
99 |
4 |
94 |
94 |
|
2500 кг (n=29 и 5) |
25 |
98 |
99 |
8 |
93 |
94 |
|
3000 кг (n=110 и 8) |
89 |
95 |
99 |
20 |
91 |
94 |
|
3250 кг (n=162 и 10) |
125 |
90 |
97 |
22 |
91 |
93 |
|
3500 кг (n=246 и 16) |
185 |
93 |
98 |
29 |
90 |
93 |
|
3750 кг (n=306 и 26) |
237 |
87 |
97 |
37 |
89 |
93 |
|
4000 кг (n=368 и 42) |
293 |
85 |
96 |
51 |
87 |
93 |
|
Искусственный стабилизирующий отбор по удою |
||||||
|
2000-5500 кг |
2 |
100 |
100 |
-15 |
96 |
94 |
|
2300-5200 кг |
3 |
100 |
100 |
-23 |
97 |
94 |
|
2600-4900 кг |
13 |
99 |
100 |
-44 |
99 |
93 |
|
2900-4600 кг |
25 |
98 |
99 |
-54 |
101 |
93 |
|
3200-4300 кг |
35 |
98 |
99 |
-74 |
105 |
94 |
|
3500-4000 кг |
45 |
91 |
99 |
-91 |
108 |
95 |
Из таблицы 6 следует, что ОПХ «Стрелецкое» группа коров-матерей с удоями менее 2500 кг молока (n=26) дала наибольшую аддитивную наследуемость удоев, процентного содержания жира в молоке и количества молочного жира – 52,4, 39,0 и 76,8% соответственно. При этом генетическая корреляция (rG) между удоем и жирностью молока составила 0,469, а дочери превзошли матерей на 1086 кг молока и 0,06% жира. От 216 матерей с удоями от 2501 до 3500 кг молока были получены дочери, которые превысили их средний показатель удоя на 299 кг молока. При закреплении к 50 матерям с удоями 4501-5500 кг молока произошёл резкий скачок генетической изменчивости жирности молока – до 56,2%. В ЗАО «Куракин-ское» ни одна группа матерей не дала существенных результатов по увеличению изменчивости.
Таблица 6 – Влияние матерей на аддитивную изменчивость и генетическую корреляцию селекционных признаков чёрно-пёстрого голштинизированного скота в хозяйствах Орловской области
|
Удой матерей за 305 дней первой лактации, кг |
Я о ч о |
Аддитивная изменчивость, % |
Фенотипические корреляции между селекционными признаками молочной продуктивности, r±m r |
1 § о |
|||||
|
Удой, кг |
Жир, % |
Жир, кг |
Удой матерей -удой дочерей |
Жир матерей -жир дочерей |
Удой матерей -жир дочерей |
Жир матерей -удой дочерей |
|||
|
ОПХ «Стрелецкое» Орловского района |
|||||||||
|
менее 2500 кг |
26 |
52,4 |
39,0 |
76,8 |
0,262±0,197 |
0,195±0,200 |
-0,241±0,198 |
0,453±0,182* |
0,469** |
|
2501-3500 кг |
216 |
29,2 |
5,4 |
9,8 |
0,146±0,067* |
0,054±0,068 |
-0,011±0,068 |
-0,002±0,068 |
-0,073 |
|
3501-4500 кг |
240 |
21,1 |
6,6 |
11,9 |
0,106 ±0,064 |
0,066 ±0,064 |
0,009 ±0,065 |
-0,039 ±0,065 |
-0,179* |
|
4501-5500 кг |
50 |
– |
56,2 |
– |
-0,199 ±0,141 |
0,281 ±0,138* |
0,213 ±0,141 |
-0,073 ±0,144 |
– |
|
ЗАО «Куракинское» Свердловского района |
|||||||||
|
менее 3000 кг |
15 |
– |
25,6 |
0,5 |
-0,072±0,277 |
0,128±0,275 |
0,039±0,277 |
0,280±0,266 |
– |
|
3001-4000 кг |
39 |
– |
– |
– |
-0,053±0,164 |
-0,211±0,161 |
-0,109±0,163 |
0,231±0,160 |
– |
|
4001-5000 кг |
122 |
– |
– |
– |
-0,099±0,090 |
-0,017±0,091 |
-0,037±0,091 |
-0,064±0,090 |
– |
|
5001 и выше кг |
37 |
1,4 |
– |
0,2 |
0,070±0,169 |
-0,112±0,168 |
0,056±0,169 |
-0,054±0,169 |
– |
Таблица 7 – Влияние процента генов голштинской породы (HF) на эффективность селекции по молочной продуктивности в ЗАО «Куракинское»
|
%HF |
n |
Аддитивная изменчивость признаков , % |
и С/) |
Относительно к матерям, % |
Относительно к прогнозу, % |
r G |
||
|
Удой, кг |
Жир, % |
Жир, кг |
||||||
|
0 |
13 |
41,8 |
28,0 |
30,4 |
57 |
96 |
103 |
0,097 |
|
25,0 |
33 |
73,4 |
– |
48,4 |
24 |
92 |
95 |
– |
|
37,5 |
28 |
49,4 |
0,4 |
63,2 |
-8 |
95 |
94 |
0,138 |
|
50,0 |
110 |
– |
– |
– |
-8 |
95 |
94 |
– |
|
62,5 |
41 |
39,0 |
– |
27,8 |
7 |
92 |
93 |
– |
|
Кроме F 1 |
105 |
54,6 |
– |
47,8 |
9 |
93 |
93 |
– |
Однако в ЗАО «Куракинское» (см. таблицу 7) прослеживалось влияние процента генов голштинской породы на аддитивную изменчивость. Так, по удою и количеству молочного жира она составила от 39,0 до 73,4% и от 27,8 до 63,2% соответственно. Наиболее удачным по генетической корреляции между удоем и жирностью молока оказался генотип с 37,5% генов голштинской породы. У чистопородных коров 3% от величины удоев составили непрогнозируемые эффекты, а в первом поколении, судя по всему, гетерозис не проявляется по причине адаптации помесей.
Кроме того, дочери быка-производителя Рома 1675 превзошли матерей на 8,1% (это можно считать относительным эффектом гетерозиса), а прогноз удоя через аддитивную наследуемость – на 3,4%, в то время как h2А удоя его дочерей составляла 12,0%. Следовательно, Ром 1675 способствовал проявлению неаддитивной наследуемости в стаде.
Таблица 8 – Эффект селекции по удою и коррелятивные сдвиги жирности молока и живой массы по первой лактации при разных моделях отбора чёрно-пёстрого голштинизированного скота в ОПХ «Стрелецкое»
|
К |
я м о ч |
''Ja |
о |
Модель отбора |
|||||||||||||||
|
По удою (I) |
По живой массе (II) |
По КМЖ/100 (III) |
По I от. (IV) |
||||||||||||||||
|
С/) |
GO |
р^ |
С/) |
С/) |
р^ |
С/) |
pf С/) |
р^ |
р^ |
С/) |
С/) |
р^ |
|||||||
|
0 |
38 |
– |
3296 |
157 |
– |
+0,05 |
-2 |
20 |
– |
+0,02 |
+5 |
157 |
– |
+0,05 |
-1 |
157 |
– |
+0,05 |
-2 |
|
12,5 |
20 |
50,2 |
4027 |
132 |
66,3 |
+0,02 |
0 |
1 |
0,50 |
0 |
+1 |
133 |
66,8 |
+0,03 |
0 |
133 |
66,8 |
+0,03 |
0 |
|
25,0 |
109 |
33,0 |
3313 |
127 |
41,9 |
+0,02 |
-1 |
8 |
2,64 |
0 |
+2 |
118 |
38,9 |
+0,02 |
+3 |
122 |
40,3 |
+0,01 |
+3 |
|
37,5 |
46 |
15,1 |
3588 |
131 |
19,8 |
+0,01 |
-1 |
-8 |
-1,21 |
+0,01 |
+6 |
130 |
19,6 |
+0,02 |
0 |
108 |
16,3 |
+0,06 |
+2,2 |
|
50,0 |
146 |
10,4 |
3538 |
132 |
13,7 |
+0,01 |
+2 |
34 |
3,54 |
0 |
+5 |
121 |
12,6 |
+0,03 |
0 |
124 |
12,9 |
+0,03 |
+2,2 |
|
62,5 |
78 |
44,6 |
3887 |
112 |
52,2 |
+0,01 |
0 |
-3 |
-1,34 |
+0,02 |
+4 |
85 |
37,9 |
+0,04 |
0 |
110 |
49,1 |
+0,04 |
+3,2 |
|
75,0 |
128 |
53,6 |
3632 |
143 |
76,6 |
+0,03 |
0 |
55 |
29,48 |
+0,03 |
+4 |
75 |
40,2 |
+0,03 |
-1 |
136 |
72,9 |
+0,04 |
+1,4 |
|
87,5 |
46 |
25,2 |
3595 |
131 |
33,0 |
+0,02 |
+6 |
4 |
1,01 |
+0,01 |
+6 |
124 |
31,2 |
+0,03 |
+2 |
131 |
33,0 |
+0,02 |
+6,2 |
|
Все |
661 |
19,1 |
3652 |
131 |
25,0 |
+0,021 |
+0,55 |
18 |
3,44 |
+0,011 |
+4,22 |
118 |
22,5 |
+0,031 |
0 |
128 |
24,4 |
+0,034 |
+2,233 |
Примечание: HF – % генов чёрно-пёстрой голштинской породы.
Из таблицы 8 следует, что при выбраковке 10% коров и учёте аддитивной генетической изменчивости для каждой группы отдельно отбор по удою в ОПХ «Стрелецкое» имел разную эффективность, а наибольший эффект селекции был характерен для помесных коров с 75% генов голштинской породы – SE=76,6 кг. Отбор по живой массе практически не вёл к увеличению удоев при разведении коров любого генотипа – 3,34 кг молока в среднем по всему стаду, хотя у коров с 75% генов голштинской породы Sd удоя составил 55 кг молока. Отбор коров, лучших по количеству молочного жира на 100 кг живой массы, давал меньший результат по всему поголовью, чем при отборе по удою (22,5 и 25,0 кг молока соответственно), в то время как по большинству генотипов в сравнении с обычным отбором по удою различия были несущественными. В целом, отбор по индексу Iот. дал больше возможностей для отбора коров, лучших одновременно по удою, жирности молока и живой массе. Это подтвердилось полученным нами эффектом селекции при отборе – 24,4 кг, а SE удоев при отборе по индексу были незначительно ниже, чем при одностороннем отборе. Повышение интенсивности отбора способствовало увеличению эффекта селекции по удою. При отборе по удою в стаде чёрно-пёстрых коров коррелятивные сдвиги по жирности молока и живой массе составили 0,021% и 0,555 кг соответственно, причём в каждой группе результативность отбора была разной. Отбор по живой массе способствовал увеличению жира и живой массы на 0,011% и 4,222 кг соответственно. Третий вариант отбора не давал увеличения живой массы, но вел к положительному коррелятивному сдвигу жирности молока – 0,031%, что было выше, чем при отборе по удою, всего на 0,01% жира. Четвёртая модель отбора способствовала одновременному увеличению жирности молока и живой массы, причём сдвиг по жирности молока являлся наибольшим среди всех вариантов отбора (0,034%), а увеличение по живой массе уступало второй модели отбора всего 1,989 кг.
Впоследствии предпринятый индексный отбор вёл к наибольшей дополнительной прибыли за счёт эффекта селекции по удою, коррелятивных сдвигов по жирности молока и живой массе коров, а следовательно, к максимальной экономической эффективности – 139965,3 руб., что было больше, чем при других моделях отбора, на 2007,4-109562,3 руб. При комплексной селекции по промерам и продуктивному индексу с увеличением выбраковки от 10 до 50% рентабельность производства молока в стаде коров повысилась с 24,6 до 40,9%. Дополнительный удой за счёт отбора по промерам увеличился с 107 до 480 кг молока. Разработанная модель отбора способствовала повышению всех экономических показателей.
Sd удоя, кг SE удоя, R жира, % R массы, Прибыль, кг кг руб
Параметры
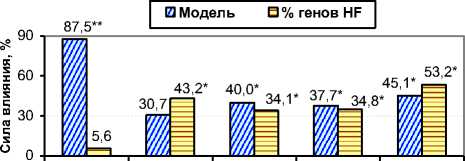
Рис. 5. Сила влияния модели отбора и % генов голштинской породы на эффективность селекции чёрнопёстрого скота в ОПХ «Стрелецкое», % (n=661).
Дисперсионный анализ показал, что модель отбора существенно влияла на селекционный дифференциал по удоям, коррелятивные сдвиги жирности молока и живой массы, а также дополнительную прибыль – h2 x =87,5% (α<1%), 40,0, 37,7 и 45,1% (α<5%) соответственно (см. рис. 5). Процент генов голштинской породы имел высокую силу влияния на большинство генетикостатистических параметров и дополнительную прибыль при производстве молока – h2x =34,1-53,2% (α<5%), за исключением селекционного дифференциала (5,6%).
Второй этап исследований показал, что при увеличении интенсивности отбора от 10 до 90% удои за 305 дней первой лактации в модельных группах возрастали от 3609 до 5043 кг молока, жирность молока – от 3,91 до 4,03%. Проявлялась тенденция к уменьшению стандартного отклонения (σ) от 746,5 до 532,4 кг молока, хотя при 70% выбраковки σ=514,5 кг молока (см. таблицу 9).
Схожие закономерности проявлялись по коэффициенту вариации удоев и количества молочного жира, однако показатели изменчивости жирности молока были стабильными (Cv=6,96-7,58% при σ=0,28-0,30%). Интенсивность отбора при этом по трём селекционным признакам возрастала с 0,179σ, 0,069σ и 0,163σ до 1,921σ, 0,414σ и 1,687σ, селекционный дифференциал – с 134 кг, 0,02% и 5,70 кг до 1434 кг, 0,12% и 58,7 кг соответственно. При этом эффект селекции по удою увеличивался линейно и при 90% составил 519,1 кг молока и 18,20 кг молочного жира, хотя за счёт аддитивного действия генов рост жирности молока составлял всего 0,01-0,02%.
Существенного внимания заслуживал тот факт, что по всему поголовью (n=542) не было превосходства над матерями по селекционным признакам (-17 кг мо- лока, -0,02% жира и -1,8 кг молочного жира), однако у 10% лучших коров (90% выбраковки) превосходство над матерями составило 1255 кг молока, 0,10% жира и 50,9 кг молочного жира.
Графическое изображение коэффициентов наследуемости показало, что при увеличении интенсивности выбраковки от 10 до 90% чёткой тенденции по генотипической изменчивости удоев получено не было. Однако её резкое возрастание в последней группе может быть подтверждением наличия непрогнозируемых генетических эффектов (доминирования, эпистаза и пр.), вызванных не только влиянием лучших генотипов матерей, но и их совместным действием с генотипами отцов.
Изучение структуры корреляций между удоями и жирностью молока показало, что независимо от группы выбраковки фенотипическая связь была положительной и составила в пределах 0,072-0,253. Схожие тенденции проявлялись по паратипической корреляции, однако генетическая корреляция между удоем и жирностью молока не проявляла стабильности в зависимости от выбраковки (от +0,434 при 50% и -0,818 при 80% выбраковки). Это может быть подтверждением скрытых генетических процессов, а также того, что генетическая корреляция между селекционными признаками зависела не только от аддитивного действия генов.
Структура корреляций между удоем и количеством молочного жира была стабильной, а сами коэффициенты в отдельных вариантах приближались к 0,85-0,98. Это подтвердило возможность увеличения селекционных признаков не только за счёт отбора, но и за счёт изменения условий кормления и содержания.
Фенотипические и генотипические корреляции между жирностью молока и количеством молочного жира были низкими или отрицательными, однако паратипические коэффициенты между данными признаками были достаточно высокими (0,525-0,579).
Удой, кг Жир, %
Интенсивность выбраковки, %
Рис. 6. Множественный плейотропный эффект по селекционным признакам, %
Таблица 9 – Эффективность одностороннего отбора чёрно-пёстрых коров по удою первой лактации в ОПХ «Стрелецкое» Орловской области
|
Признаки и параметры |
интенсивность выбраковки по удою за 305 дней лактации, % |
|||||||||
|
0 |
10 |
20 |
30 |
40 |
50 |
60 |
70 |
80 |
90 |
|
|
голов, n |
542 |
489 |
434 |
379 |
326 |
271 |
217 |
163 |
108 |
54 |
|
Продуктивность в стаде |
||||||||||
|
Удой за 305 дней, кг |
3609 |
3743 |
3852 |
3961 |
4070 |
4189 |
4326 |
4484 |
4680 |
5043 |
|
Жир, % |
3,91 |
3,93 |
3,94 |
3,95 |
3,96 |
3,99 |
4,01 |
4,02 |
4,02 |
4,03 |
|
Жир, кг |
141,1 |
146,8 |
151,4 |
156,0 |
161,0 |
166,5 |
172,7 |
179,1 |
186,6 |
199,8 |
|
Среднеквадратическое отклонение, σ |
||||||||||
|
Удой за 305 дней, кг |
746,5 |
661,9 |
619,1 |
586,0 |
561,6 |
541,7 |
524,2 |
514,5 |
524,8 |
532,4 |
|
Жир, % |
0,29 |
0,29 |
0,29 |
0,29 |
0,30 |
0,30 |
0,29 |
0,29 |
0,28 |
0,30 |
|
Жир, кг |
34,8 |
31,9 |
30,7 |
29,9 |
28,6 |
28,0 |
27,5 |
28,3 |
30,9 |
38,0 |
|
Коэффициент вариации, C v (%) |
||||||||||
|
Удой за 305 дней, кг |
20,7 |
17,9 |
16,1 |
14,8 |
13,8 |
12,9 |
12,1 |
11,5 |
11,2 |
10,6 |
|
Жир, % |
7,41 |
7,38 |
7,36 |
7,34 |
7,58 |
7,52 |
7,23 |
7,21 |
6,96 |
7,44 |
|
Жир, кг |
24,7 |
21,7 |
20,3 |
19,2 |
17,8 |
16,8 |
15,9 |
15,8 |
16,6 |
19,0 |
|
Ошибка среднего значения, ±m |
||||||||||
|
Удой за 305 дней, кг |
±32 |
±30 |
±30 |
±30 |
±31 |
±33 |
±36 |
±40 |
±51 |
±72 |
|
Жир, % |
±0,01 |
±0,01 |
±0,01 |
±0,01 |
±0,02 |
±0,02 |
±0,02 |
±0,02 |
±0,03 |
±0,04 |
|
Жир, кг |
±1,5 |
±1,5 |
±1,5 |
±1,5 |
±1,6 |
±1,7 |
±1,9 |
±2,2 |
±3,0 |
±5,2 |
|
Интенсивность отбора, i |
||||||||||
|
Удой за 305 дней, кг |
- |
0,179σ |
0,325σ |
0,470σ |
0,617σ |
0,771σ |
0,960σ |
1,171σ |
1,440σ |
1,921σ |
|
Жир, % |
- |
0,069σ |
0,103σ |
0,138σ |
0,172σ |
0,276σ |
0,345σ |
0,379σ |
0,379σ |
0,414σ |
|
Жир, кг |
- |
0,163σ |
0,296σ |
0,428σ |
0,572σ |
0,730σ |
0,908σ |
1,092σ |
1,307σ |
1,687σ |
|
Селекционный дифф |
еренциал, Sd |
|||||||||
|
Удой за 305 дней, кг |
- |
134 |
243 |
351 |
461 |
580 |
717 |
875 |
1075 |
1434 |
|
Жир, % |
- |
0,02 |
0,03 |
0,04 |
0,05 |
0,08 |
0,10 |
0,11 |
0,11 |
0,12 |
|
Жир, кг |
- |
5,70 |
10,3 |
14,9 |
19,9 |
25,4 |
31,6 |
38,0 |
45,5 |
58,7 |
|
Эффект селекции, SE |
||||||||||
|
Удой за 305 дней, кг |
- |
48,5 |
88,0 |
127,1 |
166,9 |
210,0 |
259,5 |
316,7 |
389,1 |
519,1 |
|
Жир, % |
- |
≈0 |
0,01 |
0,01 |
0,01 |
0,01 |
0,02 |
0,02 |
0,02 |
0,02 |
|
Жир, кг |
- |
1,77 |
3,19 |
4,62 |
6,17 |
7,87 |
9,80 |
11,78 |
14,11 |
18,20 |
|
Превосходство над матерями |
||||||||||
|
Удой за 305 дней, кг |
-17 |
+85 |
+191 |
+271 |
+369 |
+459 |
+599 |
+684 |
+807 |
+1255 |
|
Жир, % |
-0,02 |
-0,01 |
+0,01 |
+0,01 |
+0,02 |
+0,04 |
+0,08 |
+0,09 |
+0,08 |
+0,10 |
|
Жир, кг |
-1,8 |
+2,6 |
+7,3 |
+10,6 |
+15,2 |
+19,5 |
+26,1 |
+29,6 |
+33,6 |
+50,9 |
|
Коэффициент наследуемости в группах, с учётом выбраковки, h 2 |
||||||||||
|
Удой за 305 дней, кг |
0,362 |
0,286 |
0,350 |
0,292 |
0,300 |
0,252 |
0,366 |
0,208 |
0,022 |
0,546 |
|
Жир, % |
0,160 |
0,122 |
0,136 |
0,186 |
0,156 |
0,152 |
0,196 |
0,224 |
0,090 |
≈0 |
|
Жир, кг |
0,310 |
0,228 |
0,280 |
0,246 |
0,244 |
0,198 |
0,308 |
0,252 |
0,134 |
0,420 |
|
Коэффициент детерминации фенотипа генотипом, h = h 2 |
||||||||||
|
Удой за 305 дней, кг |
0,602 |
0,535 |
0,592 |
0,540 |
0,548 |
0,502 |
0,580 |
0,456 |
0,148 |
0,739 |
|
Жир, % |
0,400 |
0,349 |
0,369 |
0,431 |
0,395 |
0,390 |
0,443 |
0,473 |
0,300 |
≈0 |
|
Жир, кг |
0,557 |
0,477 |
0,529 |
0,496 |
0,494 |
0,445 |
0,555 |
0,502 |
0,366 |
0,648 |
|
Структура кор |
реляции между удоем и жирностью молока |
|||||||||
|
r P1P1 |
0,072 |
0,253 |
0,236 |
0,218 |
0,197 |
0,143 |
0,086 |
0,080 |
0,130 |
0,227 |
|
rA1A1 |
-0,029 |
0,294 |
-0,229 |
0,241 |
0,244 |
0,434 |
0,173 |
-0,171 |
-0,818 |
- |
|
rE1E1 |
0,108 |
0,250 |
0,382 |
0,213 |
0,187 |
0,073 |
0,057 |
0,149 |
0,176 |
- |
|
Ст |
уктура корреляции между удоем и количеством молочного жира |
|||||||||
|
r P1P1 |
0,906 |
0,877 |
0,851 |
0,825 |
0,808 |
0,769 |
0,721 |
0,670 |
0,624 |
0,526 |
|
r A1A1 |
0,764 |
0,928 |
0,930 |
0,895 |
0,885 |
0,845 |
0,866 |
0,770 |
0,407 |
0,824 |
|
r E1E1 |
0,979 |
0,862 |
0,818 |
0,801 |
0,781 |
0,749 |
0,652 |
0,641 |
0,654 |
0,513 |
|
Структура корреляции между жирностью молока и количеством молочного жира |
||||||||||
|
r P1P1 |
0,029 |
-0,010 |
0,054 |
0,050 |
0,056 |
0,051 |
0,086 |
0,097 |
0,060 |
0,089 |
|
r A1A1 |
0,035 |
0,029 |
-0,020 |
0,004 |
0,002 |
-0,010 |
0,021 |
0,052 |
0,038 |
-0,023 |
|
rE1E1 |
0,561 |
0,552 |
0,560 |
0,562 |
0,579 |
0,558 |
0,528 |
0,525 |
0,526 |
0,544 |
|
Дополнительная прибыль от производства молока на 1 голову |
||||||||||
|
Рублей |
- |
2112 |
3765 |
5421 |
7084 |
9191 |
11426 |
13820 |
16601 |
21929 |
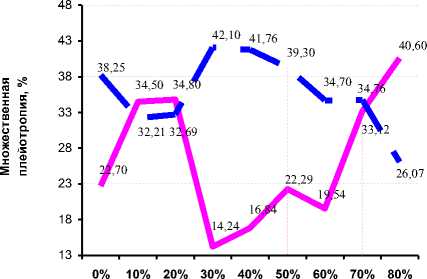
Плейотропное действие генов, кодирующих удой, на жирность молока и количество молочного жира при отсутствии выбраковки в стаде составило 22,70%, при 10 и 20% выбраковки – 34,50 и 34,80% соответственно (см. рис. 6). От 30 до 60% выбраковки прослеживался невысокий множественный плейотропный эффект, в то время как при 80% выбраковки данный эффект составил 40,60%. При 90% выбраковки в связи с отсутствием аддитивной генетической изменчивости плейотропный эффект вычислить было невозможно, хотя нельзя ис- ключать, что в незначительной степени он зависел от неаддитивных генетических факторов при 80% выбраковки. Множественный плейотропный эффект генов, отвечающих за жирность молока, был противоположным плейотропному эффекту генов, кодирующих удой.
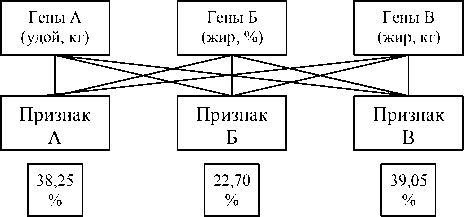
Рис. 7. Графическое изображение множественного плейотропного эффекта по признакам молочной продуктивности (n=542)
Наши исследования подтвердили наличие сложных генетических процессов при подборе и закреплении быков-производителей к стадам. Сила увеличения удоев в группах при разных вариантах подбора часто была слабо прогнозируемой, при этом точность прогноза по смежным признакам молочной продуктивности могла быть ещё более затруднительной. В этой ситуации интересным стало изучение генетико-статистических параметров при моделировании подбора.
Подтверждением того, что генетическая корреляция между удоем и жирностью молока у коров может проявляться не только за счёт матерей (т. е. аддитивной наследуемости), служат данные таблицы 10. Из их анализа следует, что матери отцов способствовали проявлению одновременного увеличения удоев и жирности молока в стаде (rG=0,902). Подобная величина генетической корреляции может быть следствием проявления комплексных генетических явлений, в том числе неаддитивной наследуемости. Кроме того, в группе коров-рекордисток влияние на одновременное увеличение
Плейотропный эффект по количеству молочного жира несущественно отличался от эффекта по жирности молока, однако был выше его при интенсивности выбраковки от 30 до 70%.
Графически множественный эффект генов, кодирующих продуктивные количественные признаки, можно представить рисунком 7, откуда следует, что около 38,25% генов, кодирующих удой, также детерминировали % жира в молоке и количество молочного жира; 22,70% генов, кодирующих жирность молока, детерми- нировали удой и количество молочного жира, аналогично 39,05% генов, кодирующих количество молочного жира, отвечали за удой и % жира в молоке.
двух признаков оказывали как матери, так и матери отцов (rG=0,477 и 0,994 соответственно). В группах про-бандов с удоями ниже 5000 кг молока за 305 дней первой лактации генетической корреляции между удоем и жирностью молока не было обнаружено. Следовательно, при подборе на одновременное увеличение двух признаков большое влияние оказывал правильный подбор лучших родительских пар.
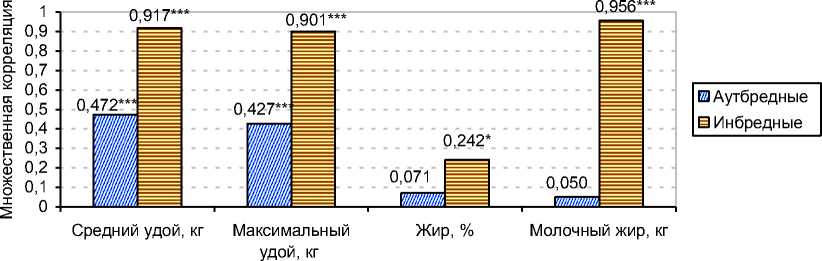
Проведённые исследования (см. рисунок 8) позволили сделать вывод, что у инбредных коров (F x =0,7812-25%) коэффициенты множественных корреляций между признаками матерей матерей, матерей и пробандов стада были существенно выше, чем в группе аутбредных коров, по величине средних удов за несколько лактаций, максимальным удоям, жирности молока и количеству молочного жира на 0,445, 0,474, 0,171 и 0,906 соответственно. Это могло быть следствием возрастания гомозиготности и увеличением аддитивной генотипической изменчивости.
Таблица 10 – Влияние материнских предков на генетические корреляции между удоем и жирностью молока первой лактации у чёрно-пёстрых голштинизированных коров СПК им. Мичурина Орловской области (n=338)
|
Группы пробандов с разным удоем, кг |
Предки |
Фенотипические корреляции, r |
r G |
|||
|
Удой матерей – удой дочерей |
Жир матерей – жир дочерей |
Удой матерей – жир дочерей |
Жир матерей – удой дочерей |
|||
|
Всё стадо |
Матери (М) |
-0,146 |
0,044 |
-0,064 |
0,078 |
- |
|
Матери отцов (МО) |
0,148 |
0,236 |
0,279 |
0,102 |
0,902*** |
|
|
5001-6000 кг |
Матери (М) |
0,043 |
0,080 |
-0,112 |
-0,056 |
0,477*** |
|
Матери отцов (МО) |
0,001 |
0,089 |
0,088 |
0,001 |
0,994*** |
|
|
4001-5000 кг |
Матери (М) |
-0,088 |
-0,058 |
0,606 |
-0,025 |
- |
|
Матери отцов (МО) |
-0,025 |
0,665 |
0,667 |
-0,025 |
- |
|
Признаки молочной продуктивности
Рис. 8. Множественная корреляция селекционных признаков матерей матерей с признаками матерей и дочерей в аутбредном (n=338) и инбредном поголовье (n=76) в хозяйствах Орловской области
Выводы и предложения (заключение). Подводя итог, следует отметить, что проведённые исследования позволяют сделать вывод, что в стадах чёрно-пёстрого скота при усилении интенсивности отбора (i=0,77-0,84σ) аддитивная изменчивость признаков молочной продуктивности может не только уменьшаться, но и возрастать. При этом до 50% поголовья может иметь положительную генетическую корреляцию между удоем и жирностью молока (rG=0,375) благодаря высокому аддитивному наследованию данных селекционных признаков, а при оптимальных вариантах выбраковки их генетическая корреляция может равняться 0,106-0,290 (р<0,05). При удачных сочетаниях родительских пар даже от групп коров со средними удоями за 305 дней первой лактации 2448-2588 кг молока можно получать дочерей с удоями на 1086-1656 кг выше, чем у матерей, и, в отдельных случаях, на 124145 кг молока выше, чем у сверстниц, полученных от матерей с удоями 3001-5000 кг. Стабилизирующий отбор по удою применять нецелесообразно, однако при его естественном проявлении (в случае выбраковки высокоудойных коров по причине гинекологических заболеваний, мастита и пр.) в стадах может сохраняться средняя аддитивная генетическая изменчивость по удою, жирности молока и количеству молочного жира. Фактические удои полностью не совпадают с прогнозом в связи с наличием неучтённых факторов, к которым можно отнести как неаддитивные эффекты, так и влияние средовых условий, взаимодействие «генотип-среда», взаимоусиливающее действие изменчивости, эффект гетерозиса, адаптационные процессы, болезни и пр. В этой связи возникает необходимость более гибкого подхода не только к определению структуры генетической изменчивости и динамики генетико-статистических параметров, но и к влиянию различных генетических групп на эффективность селекции и проявление различных генетических эффектов. Вопрос определения оптимального процента генов по голштинской породе, с одной стороны, можно считать уже неактуальным, однако проведение тщательного анализа позволяет подчеркнуть, что в группах коров с разными генотипами многие селекционные и биологические параметры селекции могут значительно отличаться. Это, наряду с моделями отбора, влияет на общую экономическую эффективность производства молока, а в каждом стаде реализа- ция селекционных признаков может иметь свои особенности, сопровождаясь различной динамикой возрастания или убывания генетической изменчивости и генетической корреляции, что, в свою очередь, по-разному отражается на результативности отбора. Отдельные быки-производители способны давать неаддитивные эффекты по признакам молочной продуктивности. Не исключено, что использование их спермы для оплодотворения удачно закреплённых коров в следующие лактации может давать схожий эффект, благодаря проявлению комбинационной способности генотипов. Так возникает необходимость накопления в племенных объединениях базы данных не только по результатам оценки производителей по продуктивности дочерей, но и по их индивидуальным особенностям проявления аддитивной и неаддитивной наследуемости.
Не следует исключать, что при одностороннем отборе по удою множественный плейотропный эффект существенно изменяется по мере возрастания интенсивности выбраковки от 10 до 90%, при этом действие генов, кодирующих удой, на жирность молока и количество молочного жира, противоположно плейотропному эффекту генов, кодирующих % жира, на удои и количество молочного жира. Судя по полученным данным, это является следствием того, что плейотропным действием обладают не только группы генов, но и отдельные гены, локализованные в разных хромосомах и проявляющие неаддитивные эффекты на признаки молочной продуктивности. В том числе, при инбридинге за счёт возрастания гомозиготности может существенно возрастать детерминация селекционных признаков матерями матерей (ММ) на продуктивность двух последующих поколений, что априори может изменить динамику множественного плейотропного эффекта при выбраковке от 10 до 90% в больших массивах инбредного поголовья. Исходя из полученных результатов, следует предложить приведенную далее схему управления динамикой генотипической изменчивости селекционных признаков и генетических корреляций между признаками (см. рис. 9).
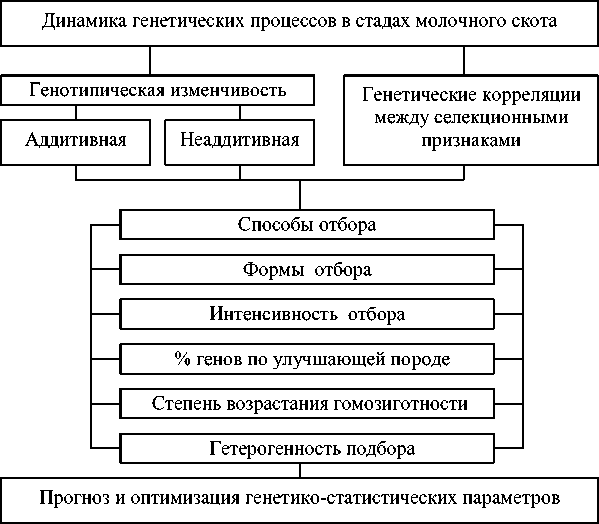
Рис. 9. Управление генетическими процессами в молочном скотоводстве
Из рисунка 9 следует, что на аддитивную и неаддитивную генотипическую изменчивость, а также на генетические корреляции между селекционными признаками можно воздействовать за счёт способов, форм и интенсивности отбора, процента генов по улучшающей породе, гетерогенности подбора и степени возрастания гомозиготности. Однако наряду с открытыми тенденциями, явлениями и закономерностями следует отметить, что фундаментальные и частные вопросы отбора и подбора в молочном скотоводстве требуют дополнительных исследо- ваний, а выбор тем кандидатских и докторских диссертаций по данному направлению должен носить приоритетный характер.
Таким образом, научно обоснованный генетикостатистический анализ должен быть основой изучения селекционного процесса в молочном скотоводстве, что позволит в дальнейшем углубленно изучить и понять закономерности динамики генетических процессов в селекции, в частности при разных способах отбора, типах, видах и вариантах подбора.
Список литературы Оценка динамики генетических процессов в молочном скотоводстве
- Айсанов З. М. Определение эффекта инбридинга у крупного рогатого скота молочных и комбинированных пород. Вестник РАСХН. 2004; 5:19-21.
- Бакай А.В., Кочиш И.И., Скрипниченко Г.Г. Генетика -М.: КолосС, 2007; 448 с.
- Винничук Д. Парадоксы инбридинга. Молочное и мясное скотоводство. 2003; 5:18-22.
- Габаев М. С., Бутырова О. А., Гукежев В. М. Эффективность разных вариантов отбора коров. Зоотехния. 2013; 5:6-7.
- Воронина Е., Стрекозов Н., Абрампальский Ф., Абылкасымов Д. Влияние вариантов подбора коров на их молочную продуктивность. Молочное и мясное скотоводство. 2007; 4:8-9.
- Дмитриев В. Д. Повышение эффективности селекции в молочном скотоводстве. Зоотехния. 2001; 4:2.
- Завертяев Б. П., Прохоренко П. Н. Совершенствование систем разведения и селекции молочного скота. Зоотехния. 2000; 8:8-12.
- Кузнецов В. М. Инбридинг в животноводстве: методы оценки и прогноза. -Киров. НИИСХ Северо-Востока, 2000; 66 стр.
- Кузнецов В. М. Разработка оптимальных программ селекции в молочном скотоводстве. Зоотехния. 1996; 1:5-13.
- Лакин Г. Ф. Биометрия. -М.: «Высшая школа», 1990; 352 с.
- Меркурьева Е. К., Абрамова З. В., Бакай А. В. и др. Генетика. -М.: ВО «Агропромиздат», 1991; 446 с.
- Мымрин В. К вопросу о гетерогенности подбора. Молочная и мясное скотоводство. 2006; 4:22-24.
- Сельцов В. И., Сермягин А. А. Продуктивные качества инбредных и аутбредных коров симментальской породы. Зоотехния. 2011; 10:2-4.
- Стародумов И. М., Гуляев С. Ю. Эффективность использования модального отбора при селекции крупного рогатого скота на молочную продуктивность. Зоотехния. 2007; 7:8-9.
- Тимофеев Ю. П., Дедов М. Д. Совершенствование методов селекции в молочном скотоводстве. Зоотехния. 1995; 4:3-6.
- Чернушенко В., Комошенков А, Бабичева В. Тип подбора родителей по ЕАВ-локусу групп крови и хозяйственно-биологические свойства дочерей. Молочное и мясное скотоводство. 2009; 2:9-10.
- Шендаков А. И. Влияние быков-производителей и типов их подбора на воспроизводительные качества чёрно-пёстрого и симментальского скота. Проблемы биологии продуктивных животных. 2011; 4 (спецвыпуск):159-162.
- Шендаков А. И. Модернизация селекции в молочном скотоводстве Орловской области. Молочное и мясное скотоводство. 2008; 6:15-19.
- Шендаков А. И. Оценка эффективности отбора скота чёрно-пёстрой породы по молочной продуктивности. Вестник ОрёлГАУ. 2010; 6:93-100.
- Шендаков А. И. Результаты комплексной оценки биологических параметров в селекции сельскохозяйственных животных. Вестник Орёл ГАУ. 2012; 6:53-63.
- Шендаков А. И., Шендакова Т. А., Ханина Т. И., Климова С. П. Повышение эффективности подбора в стадах чёрно-пёстрого и симментальского скота. Зоотехния. 2013; 3:2-6.
- Bilde T., Friberg U., Maklakov A. Fry J., Arnqvist G. The genetic architecture of fitness in a seed beetle: assessing the potential for indirect genetic benefits of choice. BMC Evolutionary Biology. 2008; 8.
- Conner J., Franks R., Stewart C. Expression of additive genetic variances and covariances for wild radish floral traits: comparison between field and greenhouse environment. Evolution. 2003; 57(3):441-451.
- Daetwyler H., Villanueva B., Bijma P., Woolliams J. Inbreeding in genome-wide selection. J. Anim. Breed. Genet. 2007; 124:369-376.
- Dal Zotto R., De Marchi M., Dalvit C., Cassandro M., Gallo L. Heritability and genetic correlations of body condition score and calving interval with yield, somatic cell score, and linear type traits in Brown Swiss cattle. J. Dairy Sci. 2007;90:5737-5743.
- Finley A. O., Banerjee S., Waldmann P., Ericsson T. Hierarchical spatial modeling of additive and dominance genetic variances for large spatial trial datasets. Biometrics. June 2009; 65:441-451.
- Parland, S. Mc., Kearney J. F., Rath M., Berry D. P. Inbreeding effect on milk production, calving performance, fertility, and conformation in Irish Holstein-Friesians. J. Dairy Science. 2007; 90:4411-4419.
- Pryce J. E., Coffey M. P., Brotherstone S. The genetic relationship between calving interval, body condition score and linear type and management traits in registered Holsteins. J. Dairy Sci. 2000; 83:2664-2671.
- Schrooten C. Genomic variation in dairy cattle -Identification and Use. Doctoral thesis (ISBN 90-8504-009-4). 2004; 69-91 (168 p.).
- Shendakov A. I. Improvement system of biological factors management in the breeding of farm animals. Russian Journal of Agricultural and Socio-Economic Sciences. 2012; 12:3-18.
- Thompson J. R., Everett R.W., Wolfe C.W. Effects of inbreeding on production and survival in Jerseys. J. Dairy Sci. 2000; 83:2131-2138.
- VanRaden P.M., Smith L.A. Selection and mating considering expected inbreeding of future progeny. J. Dairy Sci. 1999; 82:2771-2778.
- Veerkamp R.F., Brotherstone S. Genetic correlations between linear type traits, food intake, live weight and condition score in Holstein Friesian dairy cattle. Anim. Sci. 1997; 64:385-392.
- Wiggans G.R., Van Raden P.M., Zuurbier J. Calculation and use of inbreeding coefficients for genetic evaluation of United States dairy cattle. J. Dairy Sci. 1995; 78:1584-1590.
- Weigel K. A., Lin S.W. Controlling inbreeding by constraining the average relationship between parents of young bulls entering AI progeny test programs. J. Dairy Sci. 2002; 85:2376-2383.
