Особенности природы цианобактерий

Автор: Румянцев Владислав Александрович, Крюков Леонид Николаевич
Журнал: Общество. Среда. Развитие (Terra Humana) @terra-humana
Рубрика: Природная среда
Статья в выпуске: 2 (23), 2012 года.
Бесплатный доступ
Приведены сведения о последствиях цианобактериального цветения континентальных водоемов для человека, животных и гидробионтов. Рассмотрены свойства цианотоксинов. Проанализированы возможности применения биомассы цианобактерий.
Биомасса, вода, патология, применение, токсины, цветение, цианобактерии
Короткий адрес: https://sciup.org/14042712
IDR: 14042712 | УДК: 551.513
Текст научной статьи Особенности природы цианобактерий
биомасса, вода, патология, применение, токсины, цветение, цианобактерии.
К настоящему моменту выделено и охарактеризовано значительное количество цианотоксинов [39, 55]. Установлено, что аплисиатоксины и лингбиатоксин-а, являясь активаторами протеинкиназы-С, вызывают острые дерматиты, тяжелые воспаления кишечного тракта и способствуют возникновению опухолей. Липополисахариды, входящие в состав клеточной оболочки цианобактерий, вызывают раздражение кожи и аллергические реакции. Микроцистины и нодулярины относят к «факторам быстрой смерти» (гибель лабораторных животных в течение 1–4 ч). Анатоксины и сакситоксины – к «факторам очень быстрой смерти» (гибель в течение 2–30 мин). Характеристика наиболее опасных для человека цианотоксинов суммирована в табл. 1**, 3 и на рис. 1.
Сегодня при наличии набора стандартных образцов и хромато-масс-спектроскопического оборудования стало возможным идентифицировать в объектах окружающей среды весь перечень известных цианотоксинов [2; 25; 60]. Для определения токсигенности цианобактерий широко используется полимеразная цепная реакция с праймерами к генам, ответственным за синтез токсинов [28]. Быстро и надежно обнаруживать микроцистин-LR позволяют сенсоры из золотых наночастиц в углеродных нанотрубках, допированных соответствующим антителом [46]. Установлено, что микроцистины и нодулярины разрушаются озонированием, не распадаются даже после кипячения и в затененных местах водоемов сохраняются годами. Анатоксин-а(с) и сакситоксины гидролизуются. Анатоксин-а и гомоанатоксин-а подвергаются фотохимическому распаду. Деструкция цилинд-роспермозина происходит в воде при температурах свыше 50°С. Линейные изомеры микроцистинов и нодуляринов в 100 раз менее токсичны циклических структур.
Исследования продолжаются, и в 2000 г. появилось сообщение о продуцировании сакситоксина (табл. 1) пресноводной цианобактерией Planktothrix agardhii [51], колонии которых обнаружены в урбанизированных водоемах Санкт-Петербурга [7; 30].
По-видимому, еще до появления млекопитающих цианобактерии синтезировали токсины для защиты от конкурентов и обозначения сферы своего влияния. Их система адаптивного мутагенеза начала формироваться еще на заре развития жизни на Земле и за миллионы лет не раз совершенствовалась. Действительно, первые известные в ископаемом состоянии остатки организмов, найденные в осадочных породах кремния и строматолитах с возрастом 3,5–3,8 млрд лет, морфологически подобны современным нитчатым цианобактериям. Недавно строматолиты были обнаружены в подледном антарктическом озере Унтер-Зее. В наше время строматолиты (греч. «stroma» – подстилка, «litos» – камень) и онколиты (сферическая разновидность строматолитов) формируются сообществом цианобактерий с другими организмами, вокруг которого постепенно образуются карбонатные отложения (чаще известковые или доломитовые), а микроорганизмы частично отмирают или перемещаются наверх неорганических слоистых структур (рис. 2). Фактически, строматолиты являются продуктами жизнедеятельности альгобактериальных сообществ (матов), представляющих собой одну из самых устойчивых и сбалансированных экосистем с нулевым балансом [19].
Однако вернемся к вопросу о значительных количествах биомассы цианобактерий, образующихся в цветущих водохранилищах. Так, в Куйбышевском водохранилище на Волге она составляет 13–200 г/м3, в Цимлянском на Доне – от 37 до 280 г/м3, в Каховском на Днепре достигает 71–450 г/м3. Запасы цианобактерий в метровом слое воды Днепровского каскада водохранилищ в 1969 г. составляли 432407 т [27]. В настоящее время в условиях глобальной эвтрофикации водоемов, приводящей к
* Окончание. Начало см. № 1 за 2012 г., с. 232–238. Нумерация таблиц начинается в первой части статьи.
** См. № 1 за 2012 г., с. 233.
Cреда обитания
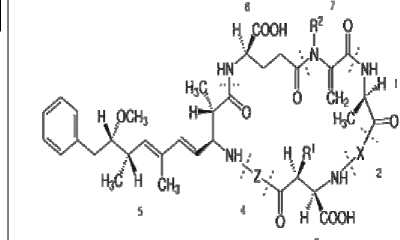
Микроцистины:
1, D-Alanine; 2, X (variable); 3, D-MeAsp (D-érythro-b-méthyl-D-acide aspartique); 4, Z (variable); 5, Adda (acide (2S, 3S, 8S, 9S)-3-amino-9-méthoxy-2-6-8-triméthyl-10-phényldéca-4,6-diènoïque); 6, D-glutamate; 7, Mdha (N-méthyldéhydroalanine). R1 et R2 sont H (déméthylmicrocystines) ou CH3.
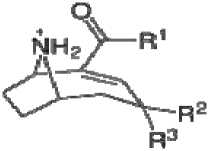
Анатоксины: анатоксин-a (R1=CH3, R2=R3=H), син-a (R1=C2H5, R2=R3=H)
гомоанаток-
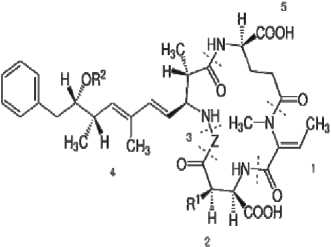
Нодулярины:
1, Mdhb (N-méthyldéhydrobutyrine); 2, -méthyl-D-acide aspartique); 3, Z (variable); 4,βD-MeAsp (D-érythro- Adda (acide (2S,3S,8S,9S)-3-amino-9-méthoxy-2-6-8-triméthyl-10-phényldéca-4,6-diènoïque); 5 D-glutamate. R1 et R2 sont H (déméthylmicrocystines) ou CH3.
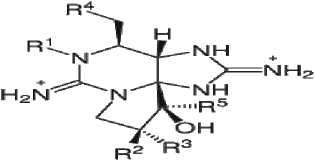
Сакситоксины:
R1=H,OH; R2=R3=H, OSO3-; R5=H,OH;
R4=OCONH2, OCOCH3, OCOC6H4, OCONHSO3
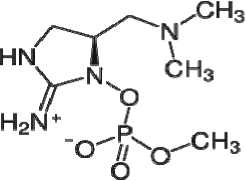
Анатоксин-a(c)
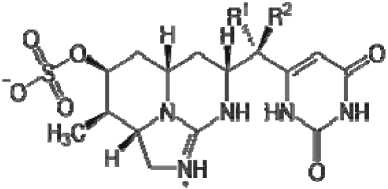
Цилиндроспермозины: R1=OH, R2=H
Рис. 1. Структурные формулы токсинов цианобактерий.
Terra Humana
Таблица 3 характеристика цианотоксинов
|
Название |
Биоактивность |
Биомишень |
Патология |
|
Микроцистины и нодулярины |
Гепатотоксичность |
Рецептор апоптоза |
Канцерогенез |
|
Анатоксины-а, а(с) и гомоанатоксин-а |
Нейротоксичность |
Ацетилхолинэстераза |
Удушье и судороги |
|
Сакситоксины |
Нейротоксичность |
Натриевые каналы |
Конвульсии и паралич |
|
Цилиндроспермо-зины |
Цитотоксичность и гепатотоксичность |
Ингибитор синтеза белка |
Некротические повреждения печени, почек, селезенки, легких, кишечника |

(a)

(b)
Рис. 2. Современные строматолиты (a) Австралии и сферическая разновидность строматолитов – онколиты (b) [].
увеличению в них запасов питательных веществ, создаются предпосылки для еще большего увеличения биомассы цианобактерий. Главным фактором, определяющим эвтрофирование, является повышение нагрузки биогенных элементов на водоем и, прежде всего фосфора, стимулирующего рост фитопланктона [53; 59]. В пределах изменения концентрации Робщ от 0,01 до 0,15 мг/л биомасса фитопланктона увеличивается за сезон от 0,4 до 20 г/м3, а содержание хлорофилла – от 1 до 65 мг/м3 [29].
Считается, что при высокой концентрации фосфора в водной среде гепатотоксич-ные штаммы цианобактерий продуцируют наибольшее количество микроцистинов [7]. Кроме того, было показано, что в цветении водоемов ведущую роль играет аминокислотная составляющая донных отложений [8]. Ввиду перечисленных обстоятельств в последние годы проводится комплексный анализ абиотических и альгологических показателей водоемов (количество биогенных веществ азота и фосфора, прозрачность воды, освещенность, глубина, содержание хлорофилла, численность и биомасса фитопланктона, наличие токсинов цианобактерий) [2]. Так, летом 2010 г. без резкого подъема концентрации биогенных элементов и в силу аномально высокой температуры воды в водохранилищах средней Волги было зафиксировано сильное цветение, вызванное массовым размножением цианобактерий Microcystis и Oscillatoria. По сравнению с обычным термическим режимом Горьковского и Чебоксарского водохранилищ содержание хлорофилла и первичная продукция фитопланктона в воде увеличились более чем в 3 раза [17]. Если прогнозы глобального потепления окажутся верными, то впереди следует ожидать значительного ухудшения экологической ситуации. Естественно, что проводятся многочисленные изыскания по улучшению качества воды и нормализации процессов регуляции в природных биоценозах цветущих водоемов [16]. Однако существенных успехов так и не достигнуто.
Ранее было замечено, что биомасса цианобактерий легко подвержена процессам брожения. Причем, применение коагулянтов (сернокислый алюминий и его производные) не останавливает брожения и тем самым открывает определенные перспективы сгущения, консервации и транспортировки сырой массы цианобактерий к местам технической переработки. Для сбора биопленок в цветущем водоеме были апробированы насосы низкого напора, предназначенные для перекачивания взвешенных веществ. Концентрация биомассы цианобактерий и сестона в собранной пульпе достигала 160–180 кг/м3 [27]. Использование распылительных сушилок для быстрого высушивания этой пульпы повышала ценность сырья без потери технологического качества. Сухая смесь может быть использована в качестве удобрений в сельском хозяйстве или для других целей, речь о которых пойдет позже. Преимущества биомассы цианобактерий перед навозом как популярным органическим удобрением отражены в табл. 4.
Экспериментально показано, что в сравнении с другими удобрениями внесение в почву биомассы цианобактерий, собранной в период цветения воды, более существенно влияет на рост, развитие и продуктивность озимой пшеницы (прирост до 18%), капусты (до 30%) и томатов (до 20%). При этом необходимо отметить, что при использовании самой разнообразной биомассы (сухой, жидкой, свежесобранной, перегнившей, представленной различным соотношением видов Microcystis, Aphanizomenon и др.) и способов ее внесения в почву не было отмечено отрицательного воздействия на жизнедеятельность и полезность опытных растений. Правда, исследователями не были приведены сведения о влиянии этих смесей на человека и сельскохозяйственных животных. В противовес сомнениям об опасности этих удобрений авторы рекомендовали использование изъятой биомассы цианобактерий без отделения от воды в
Cреда обитания
Состав природных органических удобрений
|
Удобрение |
Органические вещества, % |
Азот (N), % |
Фосфор (P), % |
Калий (K), % |
|
Навоз |
20–25 |
0,5–1,0 |
0,2–0,3 |
0,5–0,7 |
|
Биомасса цианобактерий |
до 44,0 |
2,5–8,0 |
0,7–1,2 |
0,6 |
Terra Humana
роли концентрированного раствора питательных веществ с высоким содержанием азота, взамен аммиачной воды [27].
Брожение биомассы цианобактерий наблюдается летом в местах скопления биопленок в заливах и прибрежных зонах цветущих водоемов. В числе продуктов брожения идентифицированы углекислый газ, молочная кислота, пахучие летучие органические кислоты, ацетон и спирты. Стало понятным, почему животные не обращали внимания на эти скопления, а употребляли цветущую воду с живыми микроорганизмами и затем мучились и погибали, как это описано выше. Брожению в первую очередь подвержены углеводы и при этом происходит дезактивация ирри-тантных свойств липопосахаридных токсинов цианобактерий [61]. Следовательно, применение в земледелии концентрированных суспензий цианобактерий взамен аммиачной воды не должно приводить к аллергиям и раздражениям кожи.
Важно заметить, что внесение в природную биомассу цианобактерий культуральных штаммов других микроорганизмов приводило к более низкому выходу продуктов брожения, чем в обычных условиях. Микробное сообщество цианобактерий лучше приспособлено к условиям окружающей среды и сохранению естественного баланса веществ. Не случайно, что в альгобак-териальных матах (биопленках), к примеру, щелочного термального Урчинского источника (Бурятия) обнаружено 12 видов цианобактерий, 4 вида диатомовых водорослей и 1 вид термофильных аноксигенных фотот-рофных бактерий Chloroflexus aurantiacus, делящих между собой биохимические функции без последствий для сообщества [5].
Природная биомасса цианобактерий после детоксикации может быть использована в качестве кормовых добавок в птицеводстве, животноводстве и рыбоводстве. В этом отношении решающую роль играет химический состав цианобактерий. В зависимости от вида этих прокариот в их органической части содержится разное количество углеводов (4–70%), липидов (2–12%) и белка (23–87%). По аминокислотному составу цианобактерии биологически полноценны и содержат в основном обычные аминокислоты. Причем в значительном количестве представлены незаменимые аминокислоты – изолейцин, тирозин, фенилаланин, валин, треонин, аргинин, гистидин и лизин. К сожалению, практически отсутствуют метионин и триптофан. Помимо незаменимых аминокислот в химическом составе цианобактерий обнаружены и другие ценные биологически активные вещества – разнообразные витамины, сульфатированные полисахариды с антитромбическим и противоопухолевым действием, полиглюканы (сорбенты токсинов и тяжелых металлов) и гамма-линолевая кислота, являющаяся прекурсором арахидоновой кислоты и простагландинов [43].
Содержание витаминов в значительной мере зависит от биологических особенностей вида цианобактерий и внешних условий их существования. По количеству бета-каротина цианобактерии уступают лишь зеленой галофильной водоросли Dunaliella salina, служащей сырьем для промышленного производства этого провитамина А. Содержание витамина В12 и его производных колеблется в пределах 6–8 мкг на 1 г сухого остатка и почти не зависит от вида цианобактерий. Кроме того, из цианобактерий были выделены рибофлавин, тиамин, пиридоксин, биотин и никотиновая кислота. В значительном количестве обнаружен витамин С. Помимо азота и фосфора цианобактерии богаты микроэлементами. Марганец, барий, титан, медь и цинк активно ими аккумулируются. В наибольшем количестве цианобактерии содержат железо, медь, марганец и особенно кобальт (в 4,5–55 раз больше, чем у растений).
Из цианобактерий были получены и внедрены в медицинскую практику новые цитостатики – апратоксин, ауридил, гек-тохлорин и лингбиябеллин [57]. Цианобактерии разных видов продуцируют широкий набор эффективных антибиотиков, так называемых цианобактеринов [52]. Носткарболин, выделенный из цианобактерий, сопоставим с эффективностью галантамина при лечении болезни Альцгеймера и других нейродегенеративных заболеваний [36]. В настоящее время промышленное производство ценных веществ в основном базируется на культуральных цианобактериях Spirulina platensis, о которых уже говорилось выше [37]. Сегодня этот вид цианобактерий выращивают в открытых и закрытых фотокультиваторах. Мировое производство Spirulina platensis постоянно растет и даже существуют проекты по их выращиванию в гигантских фермах на побережьи морей и океанов.
При рассмотрении всех аспектов природы токсичности цветущей воды нельзя не вспомнить о том, что реакционное равновесие всех процессов в водной среде зависит от состояния буферной емкости водоема [1]. То есть, аналогично тому, как в человеческом организме ведущее значение имеют белковый, гемоглобиновый (порфириновый аналог хлорофилла), би-карбонатный и фосфатный буфер. Способность этих буферных систем поддерживать постоянное значение рН (7,36–7,42) имеет определенные ограничения, нарушение которых приводит к патологиям. Нормальное функционирование как железо-порфирина гемоглобина млекопитающих, так и магнийпорфирина хлорофиллов цианобактерий определяется способностью этих молекулярных структур к реакциям электронного переноса и непосредственно связано с ионным составом и рН среды.
Интересно заметить, что значения рН воды, например, в Ладожском озере обычно изменяется в пределах от 7,1 до 8,4. Однако в периоды наиболее интенсивной фотосинтетической деятельности фитопланктона (1978 г. и 1984 г.) величина рН в поверхностном слое воды достигает 9,5 [18]. По сравнению с 1960 г. минерализация воды к 2000 г увеличилась в 1,2 раза и в среднем составила 63,7 мг/л. Между электропроводностью и общей минерализацией (ионизацией) водной массы озера существует прямая линейная зависимость. Многолетние наблюдения за электропроводностью воды Ладоги свидетельствуют об ее трендовом увеличении при отчетливых межгодовых (1978 г. и 1984 г.) колебаниях. Среднегодовое значение электропроводности водной массы возросло от 75,5 в 1968 г. до 85,7 мкS/см в 1989 г. К концу 90-х годов она достигла значения в 88,4 [23]. По-видимому, закономерно, что с 2004 г. в воде озера стали обнаруживать значительные количества цианотоксинов [60]. Буферная емкость природной воды Ладожского озера и условия к реакциям электронного переноса явно меняются в пользу размножения цианобактерий.
Выше отмечалось, что марганец, барий, титан, медь и цинк активно аккумулируются цианобактериями. Недавно нитчатые цианобактерии Plectonema boryanum UTEX 485 нашли применение при синтезе золотых, серебряных и палладиевых наночастиц [49]. Цианобактерии содержат сложную смесь альдегидов, сахаров, ненасыщенных кислот, многоатомных спиртов и полифенолов, действующих в качестве эффективных восстановителей катионов, а поверхностноактивные и хелатирующие вещества в их составе способны выступать в качестве стабилизаторов соответствующих наночастиц. Количество восстановителей и стабилизаторов наномасштабных частиц у разных цианобактерий существенно колеблется. Так, содержание бета-каротина в ряду Aph-anizomenon ,Microcystis и Anabaena растет от 398 до 608 мкг (на 1 г сухого вещества) и в той же последовательности увеличивается количество аскорбиновой кислоты (рис. 3). Особо следует подчеркнуть, что концентрация этих ингредиентов зависит не только от вида цианобактерий, но и от температуры и освещенности среды обитания [27].
Сегодня методы получения наночастиц с использованием природных субстанций, подобных цианобактериям, получили особую популярность [26]. Эти достижения современной «зеленой» химии имеют существенную значимость в рассмотрении причин развития токсичного цветения водоемов и подтверждаются соответствующим распределением крупности частиц донных отложений в цветущей и нецветущей природной воде. Только при практическом отсутствии наномасштабных частиц во взвесях были зафиксированы цианотоксины. [21].
Особого внимания заслуживают гуминовые вещества, которые в пресной воде образуют комплексы с ионами металлов и являются антикоагулянтами наночастиц. Как известно, взвеси донных отложений водоемов представляют собой смесь водных и почвенных гумусовых матриц, которые образуются в процессе постмортальных превращений живых организмов [20]. Вне зависимости от происхождения компонентами гумусовых матриц являются супра- и
|
СН3 СН3 СН3 СН3 н 3 сн3 сн3 сн3 Ан3 (a) |
н9 (b) |
Рис. 3. Структурные формулы бета-каротина (a) и аскорбиновой кислоты (b).
Cреда обитания
макромолекулы гуминовых веществ, различные олигомерные и низкомолекулярные соединения, неорганические ионы и гид-роксополимеры, а также наночастицы минералов. Путем изучения гелевых пленок, полученных отделением от почвенных агрегатов при подъеме на поверхность воды, стало возможным зафиксировать в этих матрицах частицы гуминовых веществ размером 2–4 нм, которые представляют собой фульвокислоты, а частицы размером 8–12 нм являются гуминовыми кислотами. Эти кислоты объединены в ассоциаты размером в десятки и сотни нанометров [31–33].
Избирательность переноса питательных веществ из гумусовых матриц внутрь клеток цианобактерий обеспечивается набором в наружной мембране пор определенного радиуса. По сути, наружная мембрана цианобактерий выступает в роли сита, задерживает ненужное и пропускает внутрь клетки необходимое в соответствии с генетически заданными параметрами. Иными словами, свойства наномасштабных элементов окружающей среды имеют не последнее значение в появлении токсичных вод [24].
Подводя итог, можно утверждать, что должно быть ясным: природа цианобактерий обладает редкой многогранностью, требует осторожности и одновременно таит в себе серьезный потенциал для научно-технического прогресса.
Список литературы Особенности природы цианобактерий
- Алекин О.А. Основы гидрохимии. -М.: Гидрометиздат, 1970. -444 с.
- Бабаназарова О.В., Кармайер Р., Сиделев С.И. и др. Структура фитопланктона и содержание микроцистинов в высокоэвтрофном озере Неро//Водные ресурсы. Т. 38. -2011, № 2. -С. 223-231.
- Белых О.И., Тихонова И.В., Сороковикова Е.Г. и др. Выявление токсичных MICROCYSTIS в озере Котокельское (Бурятия)//Вестник Томского государственного университета. 2010. №330. -С. 172-175.
- Биргер Т.И., Маляревская А.Я., Арсан О.М. К этиологии гаффской (юксовско-сартланской) болезни//Гидробиол. журн. Т. 9. -1973, № 2. -С. 115-126.
- Брянская А.В., Намсараев З.Б., Калашникова О.М. и др. Биогеохимические процессы в альгобактериальных матах щелочного термального Уринского источника//Микробиология. Т. 75. -2006, № 5. -С. 702-712.
- Волошко Л.Н., Плющ А.В., Титова Н.Н. Токсины цианобактерий (CYANOBACTERIA, CYANOPHYTA)//Альгология. Т. 18. -2008, № 1. -С. 3-20.
- Волошко Л.Н., Пиневич А.В., Копецкий И. и др. Продуцируемые цианобактериями токсины в период «цветения» воды в Нижнем Суздальском озере (Санкт-Петербург, Россия)//Альгология. -2010, № 2. -С. 210-223.
- Гладышев М.И., Колмаков В.И., Кравчук Е.С. и др. Прорастание акинет цианобактерий из донных отложений в эксперименте в водах «цветущего» и «нецветущего» водоемов//Доклады Академии наук. Т. 378. -2001, № 1. -С. 134-137.
- Горюнова С.В., Демина Н.С. Водоросли -продуценты токсических веществ. -М.: Наука, 1974. -256 с.
- Громов Б.В. Цианобактерии в биосфере//Соровский образовательный журнал. Биология. -1996, № 9. -С. 33-39.
- Гусева К.А. Роль сине-зеленых водорослей во внутренних водоемах//Вестник АН СССР. -1962, № 12. -С. 109-110.
- Гусев М.В., Никитина К.А. Цианобактерии. -М.: Наука, 1979. -228 с.
- Гусев М.В., Минеева Л.А. Микробиология. -М.: Академия, 2008. -464 с.
- Карапетян Н.В. Фотосистема 1 цианобактерий: организация и функции//Успехи биологической химии. -2001, т. 41. -С. 39-76.
- Кокшарова О.А. Применение методов молекулярной генетики и микробиологии в экологии и биотехнологии цианобактерий//Микробиология. Т. 79. -2010, № 6. -С. 734-747.
- Колмаков В.И. Методы предотвращения массового развития цианобактерий Microcystis aeruginosa Kutz emend. Elenk. в водных системах//Микробиология. Т. 75. -2006, № 2. -С. 149-153.
- Копылов А.И., Лазарева В.И., Минеева Н.М. и др. Влияние аномально высокой температуры воды на развитие планктонного сообщества водохранилищ средней Волги летом 2010 г//Доклады Академии наук. Т. 442. -2012, № 1. -С. 133-135.
- Ладожское озеро. Атлас. -СПб.: ИНОЗ РАН., 2002. -128 с.
- Литвинова Т.В. Новые данные по строению и составу строматолитовых построек (С. Прианабарье)//Литология и полезные ископаемые. -2009, № 4. -С. 428-437.
- Орлов Д.С. Гуминовые вещества в биосфере//Соросовский образовательный журнал. -1997, № 2. -С. 56-63.
- Поздняков Ш.Р., Крюков Л.Н., Румянцев В.А. Исследование влияния дисперсности водных взвесей на токсичность «цветения» воды Ладожского озера//Доклады академии наук. Т. 440. -2011, № 6. -С. 822-825.
- Прозоров А.А., Даниленко В.Н. Система «токсин-антитоксин» у бактерий: инструмент апоптоза или модуляторы метаболизма?//Микробиология. Т. 79. -2010, № 2. -С. 147-159.
- Расплетина Г.Ф., Сусарева О.М., Крючков А.М. Минерализация и электропроводность воды//Ладожское озеро -прошлое, настоящее, будущее/Ред. В.А. Румянцев. -СПб.: Наука, 2002. -С. 72-74.
- Румянцев В.А., Крюков Л.Н., Поздняков Ш.Р., Рыбакин В.Р. Наномасштабные элементы лимнологии//Вестник РАН. Т. 81. -2011, № 9. -С. 819-824.
- Русских Я.В., Чернова Е.Н., Некрасова Л.В. и др. Сравнение результатов определения цитотоксинов (анатоксина-А и микроцистина-RR) методом хромато-масс-спектроскопии, полученных с помощью приборов с различными типами ионных ловушек//Научное приборостроение. Т. 20. -2010, № 4. -С. 100-107.
- Семенова А.А., Хабатова В.В., Браже Н.А. и др. О применении наночастиц серебра, получение с использованием «зеленой химии», в диагностике объектов биологической природы методом гигантского комбинационного рассеяния//Доклады академии наук. Т. 440. -2011, № 6. -С. 773-777.
- Сиренко Л.А. Физиологические основы размножения синезеленых водорослей в водохранилищах. -Киев: Наукова думка, 1972. -203 с.
- Тихонова И.В., Белых О.И., Помазкина Г.В., Гладких А.С. Анализ цианобактерий озера Байкал и Усть-Илимского водохранилища на наличие гена синтеза микроцистина//Доклады АН. Т. 409. -2006, № 3. -С. 425-427.
- Трифонова И.С. Экология и сукцессия озерного фитопланктона. -Л.: Наука, 1990. -184 с.
- Трифонова И.С., Павлова О.А. Структура и сукцессия фитопланктона урбанизированных водоемов Санкт-Петербурга//Гидробиол. журн. Т. 41. -2005, № 1. -С. 3-12.
- Федотов Г.Н., Добровольский Г.В., Шоба С.А. Микрофазное расслоение супраполимерной гумусовой матрицы как процесс, формирующий наноструктурную организацию почвенных гелей//Доклады Академии наук. Т. 437. -2011, № 6. -С. 846-848.
- Федотов Г.Н., Добровольский Г.В., Рудометкина Т.Ф. Влияние микрофазного расслоения почвенных гелей на каталазную активность почв//Доклады Академии наук. Т. 438. -2011, № 6. -С. 842-845.
- Федотов Г.Н., Рудометкина Т.Ф. Влияние наноструктурной организации почвенных гелей на фракционный состав гумусовых веществ в почве//Доклады Академии наук. Т. 439. -2011, № 1. -С. 64-67.
- Шестаков С.В. Функциональная геномика цианобактерий//Вестник ВОГиС. Т. 8. -2004, № 2. -С. 67-72.
- Banner A.H. A dermatitis-producing alga in Hawaii. Preliminary report//Hawaii Med. J. -1959, V. 19. -P. 35.
- Becher P.G., Baumann H.I., Gademann K., Juttner F. The cyanobacterial alkaloid nostocarboline: An acetylcholinesterase and trypsin inhibitor//J. of Applied Phycology. -2009, V. 21(1). -P. 103-110.
- Belay A. The potential application of Spirulina (Arthrospira) as a nutritional and therapeutic supplement in health management//J. Amer. Nutr. Ass. V. 5. -2001, № 2. -P. 27-47.
- Buenz E.J., Howe C.L. Beta-methylamino-alanine (BMAA) injures hippocampal neurons in vivo//Neuro Toxicology. V. 28. -2007, № 3. -P. 702-704.
- Carmichael W.W., The cyanotoxins//Adv. Bot. Res. -1997, V. 27. -P. 211-256.
- Cockburn T.A., Cassanov J.G. Epidemiology of endemic cholera//Publ. Health Rep. -1960, V. 75. -P. 791.
- Codd G.A., Lindsay J., Young F.R. et al. From mass mortalities to management measures//Harmful Cyanobacteria. Netherlands: Springer, 2005. -P. 1-25.
- Cohen G., Reif C.B. Cutaneous sensitization to blue-green algae//J. Allergy. -1953, V. 24. -P. 452.
- Cohen Z. Chemicals from microalgae. -London: Taylor & Francis, 1999. -450 p.
- Dillenberg H.O., Dehnel M.K. Toxin waterbloom in Saskatchewan 1959//Canad. Med. Ass. J. -1960, V. 83. -P. 1151.
- Francis G. Poisonous Australian lake//Nature. -1878, V. 18. -P. 444, 11.
- Jing Zhang, Jianping Lei, Rong Pan et al. In siti assembly of gold nanoparticles on nitrogen-doped carbon nanotubes for sensitive immunosensing of microcystin-LR//Chem.Commun. -2011, V. 47. -P. 668-670.
- Jochimsen E.M., Carmichael W.W., An J.S. et al. Liver failure and death after exposure to microcystins at a haemodialysis center in Brazil//New Engl. J. Med. -1998, V. 338. -P. 873-878.
- Landers H. Weather aspects of Tallahassee polio-like outbreak//J.Florida Med.Ass. -1956, V. 43. -P. 355.
- Lengke M.F., Fleet M.E., Southam G. Biosynthesis of silver nanoparticles by filamentous cyanobacteria//Langmuir. -2007, V. 23. -P. 2694-2699.
- Mosher H.S., Fuhrman F.D., Buchwald J.D., Fisher H.G. Tarichotoxin-tetrodotoxin, a potent neurotoxin//Science. -1964, V. 144. -P. 1100.
- Pomati F., Sacchi S., Rassetti C. et al. The freshwater cyanobacterium Planktothrix sp. FP1: molecular identification and detection of paralytic shellfish poisoning toxins//J. Phycol. -2000, V. 36. -P. 553-562.
- Rezanka T., Dembitsky V.M. Metabolites produced by cyanobacteria belonging to several species of the family Nostocaceae//Folia Microbiol. -2006, V. 51. -P. 159-182.
- Schindler D.W. Evolution of phosphorus limitation in lakes//Science. -1977, V. 196. -P. 260-262.
- Skulberg O.M. Cyanobacteria/Cyanotoxin research -looking back for future: the opening lecture on the 6th ICTC, Bergen, Norway//Environ. Toxicol. -2005, V. 3. -P. 220-228.
- Sivonen K., Jones G. Cyanobacterial toxins. Toxic cyanobacteria in water -a guide to their public health consequences, monitoring and management. -London: E. & F.N. Spon, 1999. -P. 41-111.
- Steyn D.G. Poisoning of animals and human beings by algae//Afr. J. Sci. -1945, V. 41. -P. 243.
- Tan L.T. Bioaktive natural products from marine cyanobacteria for drug discovery//Phytochem. V. 68. -2007, № 7. -P. 954-979.
- Veldee M.V. Epidemiological study of suspected waterborne Gastroenteritas//Amer. J. Publ. Health. -1931, V. 21. -P. 1227.
- Vollenweider R.A. Scientific fundamentals of the eutrophication of lakes and flowing waters with particular reference to nitrogen and phosphorus as factors in eutrophication//Techn. Rep. Organiz. Econom. Cooper. Devel. (Paris). -1968, V. 27. -P. 1-250.
- Voloshko L., Kopecky J., Safronova T. et al. Toxins and other bioactive compounds produced by cyanobacteria in Lake Ladoga//Estonian Journal of Ecology. V. 57. -2008, № 2. -P. 100-110.
- Weckesser J., Drews G. Lipopolysaccharides of photosysnthetic prokaryotes//Ann. Rev. Microbiol. -1979, V. 33. -P. 215-239.
- World Health Organization: Guidelines for Drinking-water Quality. Health Criteria and other supporting information. -Geneva: World Health Organization, 1998. -273 p.
- Xie L., Xie P., Guo L. et al. Organ distribution and bioaccumulation of microcystins in freshwater fish at different trophic levels from the eutrophic lake Chaohu, China//Env. Toxicology. -2005, V. 20. -P. 293-300.
- Гусева К.А. Роль сине-зеленых водорослей во внутренних водоемах//Вестник АН СССР. -1962, № 12. -С. 109-110.
- Гусев М.В., Никитина К.А. Цианобактерии. -М.: Наука, 1979. -228 с.
- Гусев М.В., Минеева Л.А. Микробиология. -М.: Академия, 2008. -464 с.
- Карапетян Н.В. Фотосистема 1 цианобактерий: организация и функции//Успехи биологической химии. -2001, т. 41. -С. 39-76.
- Кокшарова О.А. Применение методов молекулярной генетики и микробиологии в экологии и биотехнологии цианобактерий//Микробиология. Т. 79. -2010, № 6. -С. 734-747.
- Колмаков В.И. Методы предотвращения массового развития цианобактерий Microcystis aeruginosa Kutz emend. Elenk. в водных системах//Микробиология. Т. 75. -2006, № 2. -С. 149-153.
- Копылов А.И., Лазарева В.И., Минеева Н.М. и др. Влияние аномально высокой температуры воды на развитие планктонного сообщества водохранилищ средней Волги летом 2010 г//Доклады Академии наук. Т. 442. -2012, № 1. -С. 133-135.
- Ладожское озеро. Атлас. -СПб.: ИНОЗ РАН., 2002. -128 с.
- Литвинова Т.В. Новые данные по строению и составу строматолитовых построек (С. Прианабарье)//Литология и полезные ископаемые. -2009, № 4. -С. 428-437.
- Орлов Д.С. Гуминовые вещества в биосфере//Соросовский образовательный журнал. -1997, № 2. -С. 56-63.
- Поздняков Ш.Р., Крюков Л.Н., Румянцев В.А. Исследование влияния дисперсности водных взвесей на токсичность «цветения» воды Ладожского озера//Доклады академии наук. Т. 440. -2011, № 6. -С. 822-825.
- Прозоров А.А., Даниленко В.Н. Система «токсин-антитоксин» у бактерий: инструмент апоптоза или модуляторы метаболизма?//Микробиология. Т. 79. -2010, № 2. -С. 147-159.
- Расплетина Г.Ф., Сусарева О.М., Крючков А.М. Минерализация и электропроводность воды//Ладожское озеро -прошлое, настоящее, будущее/Ред. В.А. Румянцев. -СПб.: Наука, 2002. -С. 72-74.
- Румянцев В.А., Крюков Л.Н., Поздняков Ш.Р., Рыбакин В.Р. Наномасштабные элементы лимнологии//Вестник РАН. Т. 81. -2011, № 9. -С. 819-824.
- Русских Я.В., Чернова Е.Н., Некрасова Л.В. и др. Сравнение результатов определения цитотоксинов (анатоксина-А и микроцистина-RR) методом хромато-масс-спектроскопии, полученных с помощью приборов с различными типами ионных ловушек//Научное приборостроение. Т. 20. -2010, № 4. -С. 100-107.
- Семенова А.А., Хабатова В.В., Браже Н.А. и др. О применении наночастиц серебра, получение с использованием «зеленой химии», в диагностике объектов биологической природы методом гигантского комбинационного рассеяния//Доклады академии наук. Т. 440. -2011, № 6. -С. 773-777.
- Сиренко Л.А. Физиологические основы размножения синезеленых водорослей в водохранилищах. -Киев: Наукова думка, 1972. -203 с.
- Тихонова И.В., Белых О.И., Помазкина Г.В., Гладких А.С. Анализ цианобактерий озера Байкал и Усть-Илимского водохранилища на наличие гена синтеза микроцистина//Доклады АН. Т. 409. -2006, № 3. -С. 425-427.
- Трифонова И.С. Экология и сукцессия озерного фитопланктона. -Л.: Наука, 1990. -184 с.
- Трифонова И.С., Павлова О.А. Структура и сукцессия фитопланктона урбанизированных водоемов Санкт-Петербурга//Гидробиол. журн. Т. 41. -2005, № 1. -С. 3-12.
- Федотов Г.Н., Добровольский Г.В., Шоба С.А. Микрофазное расслоение супраполимерной гумусовой матрицы как процесс, формирующий наноструктурную организацию почвенных гелей//Доклады Академии наук. Т. 437. -2011, № 6. -С. 846-848.
- Федотов Г.Н., Добровольский Г.В., Рудометкина Т.Ф. Влияние микрофазного расслоения почвенных гелей на каталазную активность почв//Доклады Академии наук. Т. 438. -2011, № 6. -С. 842-845.
- Федотов Г.Н., Рудометкина Т.Ф. Влияние наноструктурной организации почвенных гелей на фракционный состав гумусовых веществ в почве//Доклады Академии наук. Т. 439. -2011, № 1. -С. 64-67.
- Шестаков С.В. Функциональная геномика цианобактерий//Вестник ВОГиС. Т. 8. -2004, № 2. -С. 67-72.
- Banner A.H. A dermatitis-producing alga in Hawaii. Preliminary report//Hawaii Med. J. -1959, V. 19. -P. 35.
- Becher P.G., Baumann H.I., Gademann K., Juttner F. The cyanobacterial alkaloid nostocarboline: An acetylcholinesterase and trypsin inhibitor//J. of Applied Phycology. -2009, V. 21(1). -P. 103-110.
- Belay A. The potential application of Spirulina (Arthrospira) as a nutritional and therapeutic supplement in health management//J. Amer. Nutr. Ass. V. 5. -2001, № 2. -P. 27-47.
- Buenz E.J., Howe C.L. Beta-methylamino-alanine (BMAA) injures hippocampal neurons in vivo//Neuro Toxicology. V. 28. -2007, № 3. -P. 702-704.
- Carmichael W.W., The cyanotoxins//Adv. Bot. Res. -1997, V. 27. -P. 211-256.
- Cockburn T.A., Cassanov J.G. Epidemiology of endemic cholera//Publ. Health Rep. -1960, V. 75. -P. 791.
- Codd G.A., Lindsay J., Young F.R. et al. From mass mortalities to management measures//Harmful Cyanobacteria. Netherlands: Springer, 2005. -P. 1-25.
- Cohen G., Reif C.B. Cutaneous sensitization to blue-green algae//J. Allergy. -1953, V. 24. -P. 452.
- Cohen Z. Chemicals from microalgae. -London: Taylor & Francis, 1999. -450 p.
- Dillenberg H.O., Dehnel M.K. Toxin waterbloom in Saskatchewan 1959//Canad. Med. Ass. J. -1960, V. 83. -P. 1151.
- Francis G. Poisonous Australian lake//Nature. -1878, V. 18. -P. 444, 11.
- Jing Zhang, Jianping Lei, Rong Pan et al. In siti assembly of gold nanoparticles on nitrogen-doped carbon nanotubes for sensitive immunosensing of microcystin-LR//Chem.Commun. -2011, V. 47. -P. 668-670.
- Jochimsen E.M., Carmichael W.W., An J.S. et al. Liver failure and death after exposure to microcystins at a haemodialysis center in Brazil//New Engl. J. Med. -1998, V. 338. -P. 873-878.
- Landers H. Weather aspects of Tallahassee polio-like outbreak//J.Florida Med.Ass. -1956, V. 43. -P. 355.
- Lengke M.F., Fleet M.E., Southam G. Biosynthesis of silver nanoparticles by filamentous cyanobacteria//Langmuir. -2007, V. 23. -P. 2694-2699.
- Mosher H.S., Fuhrman F.D., Buchwald J.D., Fisher H.G. Tarichotoxin-tetrodotoxin, a potent neurotoxin//Science. -1964, V. 144. -P. 1100.
- Pomati F., Sacchi S., Rassetti C. et al. The freshwater cyanobacterium Planktothrix sp. FP1: molecular identification and detection of paralytic shellfish poisoning toxins//J. Phycol. -2000, V. 36. -P. 553-562.
- Rezanka T., Dembitsky V.M. Metabolites produced by cyanobacteria belonging to several species of the family Nostocaceae//Folia Microbiol. -2006, V. 51. -P. 159-182.
- Schindler D.W. Evolution of phosphorus limitation in lakes//Science. -1977, V. 196. -P. 260-262.
- Skulberg O.M. Cyanobacteria/Cyanotoxin research -looking back for future: the opening lecture on the 6th ICTC, Bergen, Norway//Environ. Toxicol. -2005, V. 3. -P. 220-228.
- Sivonen K., Jones G. Cyanobacterial toxins. Toxic cyanobacteria in water -a guide to their public health consequences, monitoring and management. -London: E. & F.N. Spon, 1999. -P. 41-111.
- Steyn D.G. Poisoning of animals and human beings by algae//Afr. J. Sci. -1945, V. 41. -P. 243.
- Tan L.T. Bioaktive natural products from marine cyanobacteria for drug discovery//Phytochem. V. 68. -2007, № 7. -P. 954-979.
- Veldee M.V. Epidemiological study of suspected waterborne Gastroenteritas//Amer. J. Publ. Health. -1931, V. 21. -P. 1227.
- Vollenweider R.A. Scientific fundamentals of the eutrophication of lakes and flowing waters with particular reference to nitrogen and phosphorus as factors in eutrophication//Techn. Rep. Organiz. Econom. Cooper. Devel. (Paris). -1968, V. 27. -P. 1-250.
- Voloshko L., Kopecky J., Safronova T. et al. Toxins and other bioactive compounds produced by cyanobacteria in Lake Ladoga//Estonian Journal of Ecology. V. 57. -2008, № 2. -P. 100-110.
- Weckesser J., Drews G. Lipopolysaccharides of photosysnthetic prokaryotes//Ann. Rev. Microbiol. -1979, V. 33. -P. 215-239.
- World Health Organization: Guidelines for Drinking-water Quality. Health Criteria and other supporting information. -Geneva: World Health Organization, 1998. -273 p.
- Xie L., Xie P., Guo L. et al. Organ distribution and bioaccumulation of microcystins in freshwater fish at different trophic levels from the eutrophic lake Chaohu, China//Env. Toxicology. -2005, V. 20. -P. 293-300.
