Ультраструктурные особенности микроспорогенеза у подсолнечника Helianthus annuus L. с цитоплазматической мужской стерильностью типа RIGO

Автор: Усатов А.В., Федоренко А.Г., Тихонова М.А., Гаврилова В.А., Маркин Н.В.
Рубрика: Селекция и семеноводство
Статья в выпуске: 2 (151-152), 2012 года.
Бесплатный доступ
Методами световой и электронной микроскопии изучен микроспорогенез фертильного и ЦМС RIG0 аналогов инбредной линии подсолнечника ВИР 109. Выявлены различия в ультраструктуре клеток тапетума и микроспороцитов, возникающие при нормальном развитии пыльцы и ее дегенерации. На основании данных литературы и собственных исследований проведен сравнительный анализ морфологических особенностей, характеризующих абортивный микроспорогенез у форм с ЦМС РЕТ1 и RIG0.
Цитоплазматическая мужская стерильность (цмс), микроспорогенез, ультраструктура, подсолнечник
Короткий адрес: https://sciup.org/142151095
IDR: 142151095 | УДК: 575.153:
The ultrastructural features of microsporogenesis in sunflower Helianthus annuus L. with RIGO type of cytoplasmic male sterility
Microsporogenesis in fertile and CMS RIG0 analogs of inbred sunflower line VIR 109 was investigated by light and electron microscopy. Differences in the ultrastructure of the tapetum cells and microsporocytes between the normal pollen development and its degeneration were detected. Based on the literature and our own research, a comparative analysis of the cytologic features that characterize the abortive microsporogenesis in forms with CMS and RET1 RIG0 was conducted.
Текст научной статьи Ультраструктурные особенности микроспорогенеза у подсолнечника Helianthus annuus L. с цитоплазматической мужской стерильностью типа RIGO
Введение. Цитоплазматическая мужская стерильность (ЦМС) растений представляет собой феномен ядерно-цитоплазматического взаимодействия, в котором участвуют как гены органелл (митохондрий), так и ядра. В связи с широким использованием ЦМС-растений в качестве материнских форм для получения промышленных гетерозисных гибридов подсолнечника исследования нарушений микроспорогенеза, возникающих при различных типах ЦМС, имеют не только фундаментальную, но и практическую ценность. В настоящее время в гетерозисной селекции подсолнечника преобладает ЦМС типа РЕТ1, открытая П. Леклерком у межвидового гибрида Helianthus petiolaris Nutt. × H. annuus L. (Leclercq, 1969) . Главные структурные признаки, характеризующие абортивное развитие микроспор у линий с ЦМС типа РЕТ1, по сравнению с фертильными аналогами, были определены рядом исследователей ранее (Horner, 1977; Симоненко, 1975, 1982; Симоненко, Карпович, 1978; Laveau et al., 1989; Smart et al., 1994; Meric et al., 2003; Усатов и др., 2010). Данный тип ЦМС начинает проявляться уже на уровне спорофита в виде аномального развития тапетальной ткани, с ее последующей гипертрофией, что приводит к полной дегенерации микроспороцитов.
Кроме того, были получены данные по микроспорогенезу у подсолнечника с еще несколькими типами ЦМС. Так, у форм с ЦМС на основе H. petiolaris fallax Heiser начало процессов дегенерации пыльцы наблюдали позже, чем у ЦМС РЕТ1, на стадии формирования эндэкзины пыльцевых зерен (Laveau et al., 1989). У линий с ЦМС RES1 (на основе H. resinosus Small) мейотическое деление микроспороцитов до стадии тетрад протекает нормально, а отсутствие фертильной пыльцы обусловлено нарушениями, возникающими на постмейотических стадиях (Echeverria et al., 2003). При ЦМС типа GIG2 (на основе H. giganteus L.) формирование микроспор нарушается непосредственно во время мейотического деления микроспороцитов, а тапетум и промежуточный слой клеток стенки пыльника аномально разрастаются, как и при ЦМС РЕТ1 (Zhang et al., 2010).
Целью нашей работы является сравнительное ультраструктурное исследование особенностей микроспорогенеза у стерильной формы подсолнечника c новым типом ЦМС – RIG0 (на основе H. rigidus Desf.) и ее фертильного аналога.
Материалы и методы. Материалом исследования служили фертильный и стерильный (ЦМС RIG0) аналоги линии культурного подсолнечника ВИР 109 (селекции Кубанской опытной станции Всероссийского института растениеводства). Для электронно-микроскопического анализа отбирали пыльники фертильных и стерильных растений на начальных (стадия спорогенных клеток и микроспороцитов) и более поздних (стадии диад и тетрад микроспор) стадиях развития.
Пыльники фиксировали в 2,5 %-ном растворе глютарового альдегида в фосфатном буфере по Миллонигу (рН 7,2– 7,4) в течение 12 часов. Затем их дофик-сировали в 2 %-ном растворе OsO 4 на том же буфере в течение 1,5 часов (Усатов и др., 2009). Далее образцы обезвоживали в серии спиртов и ацетонов, и заливали в Эпон. Полутонкие и ультратонкие срезы получали на приборе EM UC26 (Leica, Германия). Ультратонкие срезы контрастировали уранил-ацетатом и цитратом свинца и исследовали в электронном микроскопе Tecnai 12 (Phillips, Голландия). Полутонкие срезы толщиной 0,5–1 мкм для светооптического наблюдения дополнительно окрашивали метиленовым синим и исследовали в светооптическом микроскопе Axioskop 40 FL (Carl Zeiss AG, Германия).
Результаты и обсуждение. Сравнительный анализ светооптической микроскопии показал, что на ранних этапах развития пыльники ЦМС RIG0 и фертильной формы линии ВИР 109 схожи. Они состоят из четырех микроспораниев (гнезд) с четырехслойными стенками, представленными эпидермисом, эндоте-цием, промежуточным слоем и тапетумом. Внутри гнезда пыльника находятся многочисленные спорогенные клетки (рис. 1).
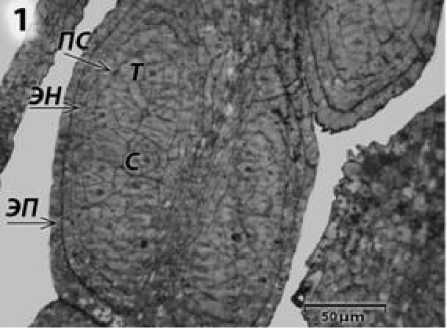
Рисунок 1 – Строение гнезда пыльника подсолнечника с ЦМС RIG0 на предмей-отической стадии развития пыльника. ЭП – эпидермис; ЭН – эндотеций; ПС – промежуточный слой клеток; Т – тапетум; С – спорогенные клетки.
Ранее нами было показано, что спорогенные клетки, локализованные в центре гнезда пыльника фертильного аналога линии ВИР 109, соединены между собой и с тапетумом плазмодесмами. Тапеталь-ные и спорогенные клетки содержат крупные ядра с хорошо заметными ядрышками (рис. 2). Эндоплазматический ретикулум (ЭПР) в клетках тапетума и микроспороцитах представлен в виде узких протяженных канальцев, на поверхности которых расположены рибосомы. (Усатов и др., 2010).
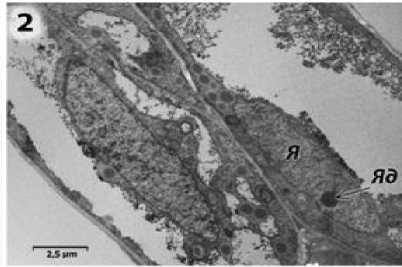
Рисунок 2 – Клетки тапетума фертильной формы подсолнечника на предмейо-тической стадии развития пыльника.
Я – ядро; Яд – ядрышко.
В цитоплазме клеток тапетума фертильной формы ВИР 109 можно наблюдать большое количество свободных рибосом, а также многочисленные митохондрии с плотным матриксом и хорошо развитыми кристами (рис. 3). Хорошо заметен аппарат Гольджи (рис. 3) и пластиды (рис. 4).
Цитоплазма микроспороцитов фертильных растений линии ВИР 109 содержит пластиды и митохондрии с большим количеством крист (рис. 5). Аппарат Гольджи не обнаружен.
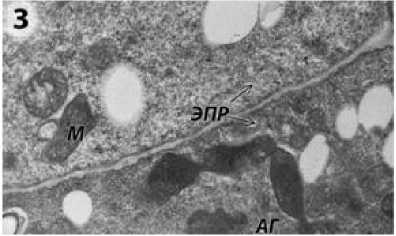
Рисунок 3 – Фрагмент клетки тапетума фертильной формы подсолнечника на предмеойтической стадии развития пыльника. эпр – эндоплазматический ретикулум;
М – митохондрии; АГ – аппарат Гольджи.
Микроспоры у фертильных растений формируются в результате двух мейоти-ческих делений спороцита и образуют сначала диады, а затем тетрады. После выхода из тетрады микроспора приобретает сферическую форму, начинает увеличиваться в размерах, ее оболочка достраивается и утолщается (Симоненко, Карпович, 1978; Усатов и др., 2010).
Клетки тапетума на стадии диад содержат по два крупных ядра (рис. 6). В цитоплазме клеток видна обширная сеть ЭПР, плеоморфные митохондрии (рис. 6, 7).
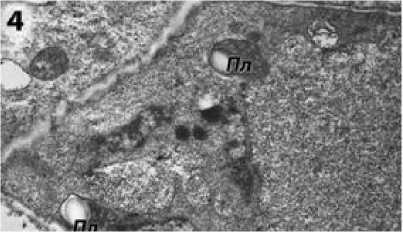
Рисунок 4 – Пластиды в клетке тапетума фертильной формы подсолнечника на предмеойтической стадии развития пыльника. Пл – пластиды.
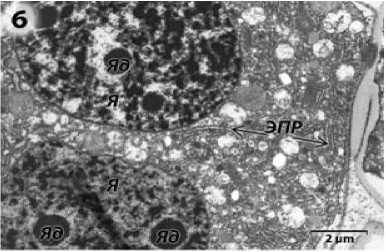
Рисунок 6 – Фрагмент клетки тапетума фертильного подсолнечника на стадии диад микроспор. я – ядра; я д – ядрышки; ЭПР – эндоплазматический ретикулум.
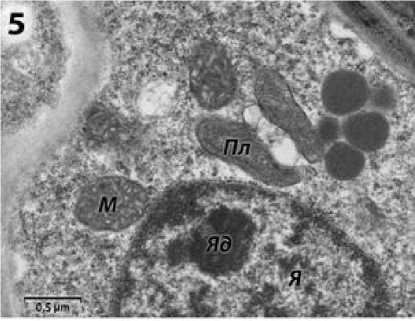
Рисунок 5 – Фрагмент спорогенной клетки фертильной формы подсолнечника на предмеойтической стадии развития пыльника. я – ядро; я д – ядрышко; м – митохондрии; Пл – пластиды.
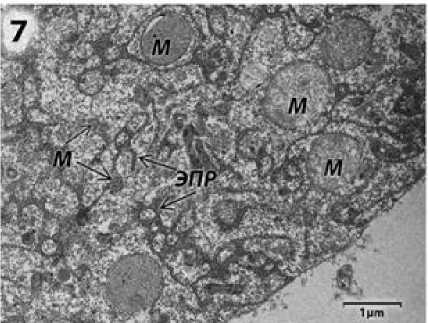
Рисунок 7 – Участок цитоплазмы клетки тапетума фертильного подсолнечника на стадии диад микроспор. м – митохондрии;
ЭПР – эндоплазматический ретикулум.
По мере роста микроспор вещество та-петального периплазмодия расходуется на питание пыльцевых зерен и формирование экзины, и к концу развития в гнездах зрелых пыльников находятся лишь трехклеточные пыльцевые зерна с характерными удлиненными спермиями и ши-пиками на поверхности экзины.
У растений линии ВИР 109 с ЦМС типа RIG0 на стадии трансформации спорогенных клеток в микроспороциты ярко выражена гипертрофия тапетальной ткани (рис. 8). Оболочки клеток тапетума исчезают (рис. 9). Гомогенный тапетум окружает спорогенный комплекс клеток, но не проникает между ними, и спороциты на протяжении всего развития пыльника имеют вид компактной массы.
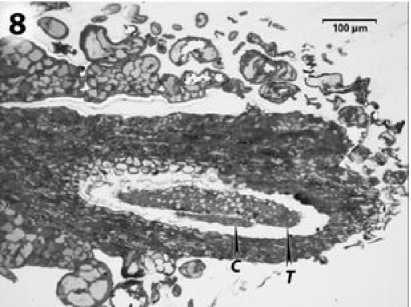
Рисунок 8 – Гнездо пыльника подсолнечника с ЦМС RIG0. т – тапетум; с – спороциты.
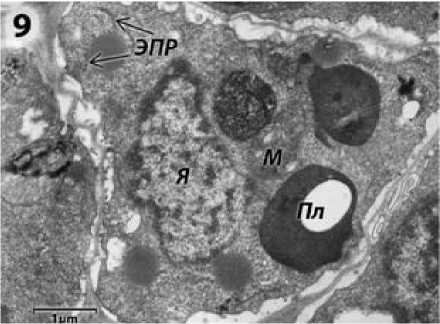
Рисунок 9 – Клетка тапетума подсолнечника с ЦМС RIG0 на стадии поздних микроспороцитов и диад. я – ядро; эпр – эндоплазматический ретикулум; М – митохондрия; Пл – пластиды.
Постепенно пространство, занимаемое спорогенным комплексом в гнезде пыльника, уменьшается, а тапетумом – увеличивается (рис. 8). Эти наблюдения согласуются с данными литературы по развитию пыльника у ЦМС РЕТ1 подсолнечника. Однако нами было показано, что у линии ВИР 109 с ЦМС РЕТ1 тапетум начинает разрастаться позже – после завершения мейотического деления микроспороцитов и образования тетрад микроспор (Усатов и др., 2010).
Цитоплазма клеток тапетума менее плотная, чем у фертильной формы, но так же содержит каналы ЭПР (их профили на срезах имеют вид замкнутых цистерн с мембранами, на поверхности которых расположены рибосомы) (рис. 9). Однако чрезмерного отложения спорополленина в виде электронноплотных полусферических «бляшек», как это наблюдал Симоненко (1982) в тапетуме растений с ЦМС РЕТ1, у растений ВИР 109 как с ЦМС RIG0, так и РЕТ1 (Усатов и др., 2010) не обнаружено. В тапетальных клетках находятся митохондрии с хорошо выраженной и мощно развитой системой крист (рис. 9). Пластиды имеют плотную строму с редкими ламеллами, форма их округлая, как и у ЦМС РЕТ1 растений линии ВИР 109 (Усатов и др., 2010), но в отличие от овальных пластид других ЦМС РЕТ1 линий (Симоненко, 1982) (рис. 9). Ядра крупные, сильно хромати-зированные, количество частиц плотного гетерохроматина внутри них чрезвычайно велико (рис. 9). Форма ядер амебоидная, с многочисленными выростами и инвагинациями. В отличие от наблюдений ЦМС РЕТ1 Симоненко (1982) ядерная оболочка у гипертрофированных амебоидных ядер линии ВИР 109 с ЦМС РЕТ1 (Усатов и др., 2010) и RIG0 не фрагментирована. Диктиосомы, в отличие от ВИР 109 с ЦМС РЕТ1 (Усатов и др., 2010), отсутствуют. В итоге, цитоплазма тапетума как ЦМС РЕТ1, так и RIG0 растений подсолнечника дегенерирует (Симоненко, 1982; Laveau et al., 1989; Усатов и др., 2010).
В спорогенной части пыльника у ВИР 109 с ЦМС RIG0, как и у линий с ЦМС РЕТ1, изученных Симоненко (1982), четкие признаки аномального развития проявляются на стадии микроспороцитов. У ВИР 109 с ЦМС РЕТ1 эти признаки начинают проявляться позже, на стадии тетрад (Усатов и др., 2010). В отдельных участках цитоплазма микроспороцитов просветляется, и возникают вакуоли, состоящие из пузырьков разного диаметра и липидных капель. В ядрах можно видеть крупные частицы конденсированного хроматина (рис. 10).
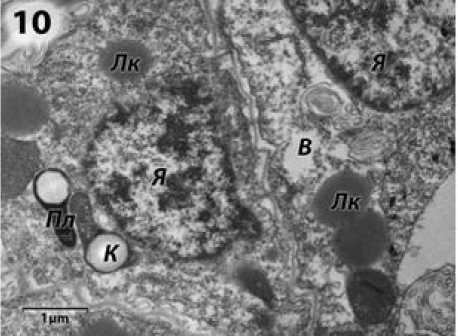
Рисунок 10 – Микроспороциты подсолнечника с ЦМС RIG0 на стадии поздних микроспороцитов и диад. я – ядро; В – вакуоли; ЛК – липидные капли; Пл – пластиды;
К – крахмал.
Большинство органелл в цитоплазме микроспороцитов набухшие, в отличие от фертильной формы, почти лишенные внутренних мембран. Пластиды округлые и гантелевидные с крупными крахмальными гранулами и едва заметными ла-меллами на периферии (рис. 10, 11), митохондрии со светлым матриксом, малочисленными кристами (рис. 11). Локализация митохондрий вблизи ядерной мембраны, как у фертильных растений, не наблюдается. Диктиосомы отсутствуют.
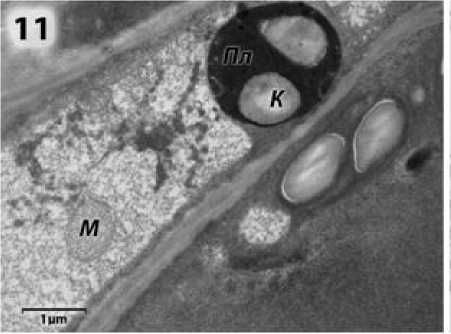
Рисунок 11 – Участок цитоплазмы микроспороцита подсолнечника с ЦМС RIG0 на стадии поздних микроспороцитов и диад: п л – пластида; к – крахмал;
М – митохондрия.
При дальнейшем развитии микроспороциты сжимаются агрессивно разросшимся тапетумом и образуют своеобразную плотную массу. Клетки ее постепенно дегенерируют, затем разрушается и все остальное содержимое гнезда пыльника. Клетки промежуточного слоя, уплощенные на предыдущих стадиях развития пыльника, сильно вакуолизируются, пузыревидно вздуваются (Симоненко, 1982), их ранее плотная цитоплазма просветляется, располагаясь пристенно (рис. 12).
Таким образом, можно заключить, что у растений подсолнечника с ЦМС типа RIG0 на ранней предмейотической стадии развития происходит интенсивное разрастание тапетальной ткани, что ведет к последующей дегенерации микроспороцитов.
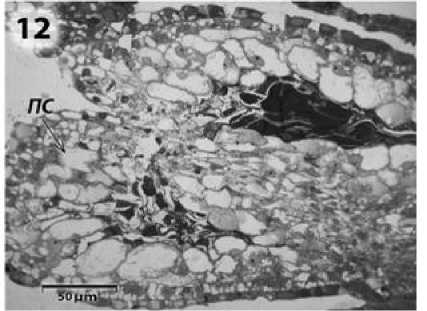
Рисунок 12 – Прекратившие развитие гнезда пыльника подсолнечника с ЦМС
RIG0. ПС – промежуточный слой клеток.
Кроме того, возникают изменения в структуре клеток тапетума (расширение каналов ЭПР, вакуолизация цитоплазмы, появление амебоидных ядер) и микроспороцитов (нарушение внутренней структуры митохондрий и пластид) стерильной формы по сравнению с фертильной. В целом процессы, выявленные нами при абортивном микроспорогенезе у растений с новым типом ЦМС RIG0, в общих чертах сходны с таковыми, описанными для ранее изученной ЦМС типа РЕТ1. Однако на исследованной нами ранней стадии развития пыльника (стадия микроспороцитов и диад) имеется ряд отличий. В частности, в клетках тапетума не происходит чрезмерного отложения спорополленина; форма тапетальных пластид округлая, а не овальная; оболочка гипертрофированных ядер тапетума на данной стадии не фрагментирована.
Выше отмеченные различия в строении ядер, пластид и митохондрий клеток тапетума и микросопороцитов у изученной нами линии ВИР 109 с новым ЦМС типа RIG0 по сравнению с фертильным и ЦМС РЕТ1 аналогами, отражают влияние митохондриального генома на протекание абортивного микроспорогенеза.
Исследование выполнено в рамках государственной темы Министерства образования и науки РФ (регистрационный номер: 4.5642.2011), при финансовой поддержке ФЦП Министерства образования и науки РФ, госконтракты № 16.740.11.0485 и № 16.552.11.7024.
